EU publishes Short-term Outlook for Agricultural Markets (Autumn 2024)
The EU’s Short-term Outlook for Agricultural Markets (Autumn 2024) reveals significant challenges in agriculture, with adverse weather, geopolitical instability, and fluctuating trade conditions impacting production. The report identifies declining cereal and oilseed outputs, particularly for soft wheat and maize. Meanwhile, milk production is expected to remain stable despite a shrinking cow herd, and the meat sector shows mixed trends, with poultry production rising but pigmeat and beef facing structural challenges.
EU’s Short-term Outlook for Autumn 2024 highlights the following key findings collectively shaping EU agricultural markets:
Weather conditions: Severe drought in Southern and Eastern Europe and excess rainfall in Northwestern regions have reduced cereal and oilseed yields.
Energy and input costs: Inflation is stabilizing but remains high, with elevated prices for inputs like fertilizers and energy.
Geopolitical tensions: The ongoing war in Ukraine and trade disputes are disrupting supply chains and impacting exports.
Global demand: While global agricultural demand is recovering, trade flows face disruptions due to regulatory changes and volatile market conditions.
Cereals
The EU cereal production in 2024/25 is projected at 260.9 million tons, approximately 7% below the 5-year average. This marks the lowest production in the past decade, driven by unfavorable weather conditions, including excessive rain in Northwestern Europe, which impacted planting, particularly for soft wheat, and drought in Southern and Eastern regions, severely affecting maize yields. Production of soft wheat and maize is expected to decline year-on-year by 9.5% and 4%, respectively. On the other hand, barley and durum wheat production are increasing by about 6% and 3%, respectively, compared to the previous year.
EU cereal exports are projected to decline by 22% year-on-year due to reduced production and quality issues. At the same time, domestic demand remains relatively stable, with animal feed consumption holding steady as livestock production stagnates. In terms of prices, cereal prices fell throughout 2024, pressuring farmers’ cash flow, which could hinder their ability to afford inputs such as fertilizers in the coming year.
Milk and Dairy Products
The EU milk market is expected to see relatively stable supply, despite a continuously shrinking cow herd. Milk yields have increased, compensating for the herd’s decline. Milk prices are forecast to stabilize after a period of volatility in the past few years, remaining above historical averages, and input costs for farmers, such as feed and energy, are showing signs of stabilizing, allowing for a potential improvement in farmer margins.
Despite the stability in milk supply, demand for dairy products continues to show mixed trends, influenced by shifts in consumer preferences and trade dynamics. The balance of milk supply and prices could provide an opportunity for dairy farmers to recover some profitability after several challenging years.
In the dairy products sector, cheese and butter continue to dominate EU production, with butter production projected to rise slightly in 2024, driven by stable milk supplies and strong domestic demand. The demand for butter in the global market remains relatively strong, although competition is rising.
Cheese production is also expected to remain stable, reflecting a balance between domestic and export markets. The cheese sector has seen steady growth over the years, supported by increasing consumer demand for premium and specialty cheeses. The demand for skimmed milk powder (SMP) and whole milk powder (WMP) is projected to remain subdued due to fluctuating global demand, particularly from key markets such as China, although some growth is expected in non-European markets.
Meat Products
The meat sector in the EU remains a mixed picture, with structural changes and external factors shaping production and trade in 2024.
Beef and Veal: Beef production continues to face structural decline due to a shrinking herd size, with the sector stabilizing but at lower levels of production. The demand for EU beef remains relatively high, and exports are increasing, but domestic production is likely to remain constrained by environmental and economic pressures. Additionally, the number of animals has been declining consistently, reflecting longer-term trends within the EU beef industry.
Pigmeat: The EU pigmeat sector is facing diverse challenges, with some countries recovering from production setbacks, while others struggle with ongoing disease outbreaks and economic issues. The overall EU pigmeat production is expected to decline slightly, and exports have become less competitive, particularly with reduced demand from key markets such as China. However, opportunities exist in other Asian countries, where EU exporters are gaining ground. Domestically, consumption is forecast to decrease slightly, reflecting shifting consumer preferences toward plant-based alternatives and poultry.
Poultry: Poultry production is expected to rise, driven by strong domestic demand and favorable export conditions. The EU poultry sector has shown resilience, with increasing production and exports, despite higher input costs. Poultry remains a preferred source of protein for consumers, especially as prices for other meats rise. The sector continues to grow in competitiveness on the international stage, with exports expected to increase in 2024 despite the challenges posed by higher EU prices.
Sheep and Goat Meat: Production of sheep and goat meat continues to decline due to the structural reduction of flocks across the EU. High EU prices have made sheep and goat meat less competitive on the global market, reducing export opportunities. Domestically, consumption remains stable but at lower levels than other meat types. The ongoing structural decline in the sector highlights long-term challenges related to animal health, productivity, and market competitiveness.
Volatility and challenges persist
The report highlights the ongoing challenges faced by the cereals, dairy, and meat sectors. Weather conditions and global trade dynamics are shaping the future of EU agriculture, with many sectors grappling with production declines and shifting market demands. Despite these challenges, opportunities exist for some areas of growth, particularly in dairy and poultry, where rising consumer demand and stable supply conditions offer optimism for the future.
Piglet rearing – there is still room for improvement!
By I. Heinzl, Editor, and Predrag Persak, Regional Technical Manager North Europe
Optimal rearing conditions for piglets are crucial for ensuring their healthy growth, reducing mortality, and enhancing productivity. These conditions include proper temperature, nutrition, housing, hygiene, and care. Here are the key aspects:
1. Temperature and ventilation
Piglets are sensitive to cold because they cannot regulate their body temperature effectively in the first few days after birth. Proper temperature control is essential to prevent chilling, possibly leading to illness and death. Additionally, regulating the temperature would cost energy, which otherwise could be spent for growth.
Signs of a too-cold environmental temperature are piling on top of one another, tucking the legs under the body, being unable to get up, laying near a corner or wall, or shivering, which may stop if the conditions worsen. Measuring the body temperature shows less than 35°C in the case of chilling.
The following temperatures are recommended for successful piglet rearing:
Farrowing unit (for newborns)
32 – 35°C (90–95°F) during the first few days
After the first week
The temperature can gradually decrease by about 1.5-2.0°C per week until it reaches 25°C (77°F)
For supplemental heating, heat lamps, heated floors, or creep areas (a designated warm spot) can be used to maintain the ideal temperature, especially in cooler climates.
Temperature is often closely related to ventilation. Ventilation is essential to reduce dust, humidity, ammonia, and other harmful substances occurring in the air. However, if fresh/cold air enters the pigsty, the temperature decreases, which can get dangerous for the piglets. Suitable ventilation means finding a good balance between providing fresh air and maintaining temperature to prevent energy losses and chilling of the piglets.
Comfort zones can be a solution. They are an effective way to keep the piglets warm and ventilation rates where needed to maintain proper air exchange and humidity levels.
2. Nutrition
Nutrition is critical for piglet growth and immune system development. Most important after birth is the access to colostrum. Piglets are born with an immature immune system, and the maternal antibodies ingested with the colostrum are vital for their survival. They should consume colostrum within the first 6 hours after birth.
It will take 5 to 7 days for piglets to stabilize and get regular on suckling schedule.
At around seven days of age, it is recommended to introduce a highly digestible, nutrient-dense creep feed that helps transition piglets from milk to solid food. Fresh and clean water of the best quality must always be available.
Never forget most important nutrient, beside sow´s love and care – water. Allow piglets free access to the excellent quality water.
3. Housing and Space
A well-designed, clean, and dry environment is critical for reducing stress and promoting health. Farrowing crates help prevent sows from accidentally crushing the piglets during the first few weeks. However, these farrowing crates should provide enough space for the sow to nurse the piglets while allowing piglets to move freely.
Separate warm and clean areas (creep spaces) for the piglets within the farrowing pen are helpful to help the piglets escape from cooler or potentially dangerous parts of the crate. Straw, sawdust, or rubber mats should be provided to keep the piglets warm and comfortable, and good drainage is essential to maintain dryness.
4. Hygiene and Health
Hygiene is crucial to prevent disease and promote the health of piglets. For this purpose, pens and farrowing units should be thoroughly cleaned. Regular removal of waste and keeping bedding dry helps control pathogens. It is essential to clean and disinfect the farrowing unit from one farrowing to the other to reduce disease risks.
Health: After birth, the piglets’ umbilical cord stump should be disinfected to prevent infections. A further essential precautionary measure to prevent anemia is an oral supplementation or an iron injection within the first three days of life, as piglets are born with low iron levels.
For further health monitoring and management, it should be ensured that the piglets are vaccinated against common diseases, such as E. coli, Mycoplasma, and Porcine Circovirus. Additionally, deworming protocols and monitoring for signs of parasites should be implemented for parasite control.
5. Weaning Practices
Piglets are typically weaned between 3 and 4 weeks of age, but early weaning (around 21 days) can be practiced in intensive systems. Optimal weaning requires gradual adaptation to solid feed and a stress-free environment.
If the piglets are weaned at 21 to 28 days, a high-quality starter diet after weaning is essential to maintain growth rates and minimize post-weaning stress.
6. Minimizing Stress
Stress management is essential to prevent disease and poor growth. For this purpose, minimize handling to the minimum during the first few days and, if necessary, handle the piglets gently to reduce stress.
A new environment also means strain for the piglets, so keep the litter groups together during weaning to reduce fighting and social stress.
7. Supportive functional feed ingredients
Depending on veterinary and managing practices, the availability of feed, and the possible use of antimicrobials or other medicals as prophylactics, there can be high variability in rearing conditions in diverse areas of the world. In the following, two functional feed ingredients with entirely different modes of action are presented that support piglets at different rearing conditions.
7.1 Egg immunoglobulins (IgY) support piglets under poor rearing conditions
Egg immunoglobulins are beneficial if piglets are not raised under the best conditions, meaning lower hygienic standards and higher pathogenic pressure. With egg immunoglobulins coming from hens having been in contact with pathogens relevant to piglets, it is possible to support the young animals. What is the background? Hens are able to transfer maternal antibodies against diseases that they are confronted with to the egg. With this mechanism, they can provide their progeny with a starter kit for the first time after hatching. However, the best thing is that these antibodies are also helpful for mammals.
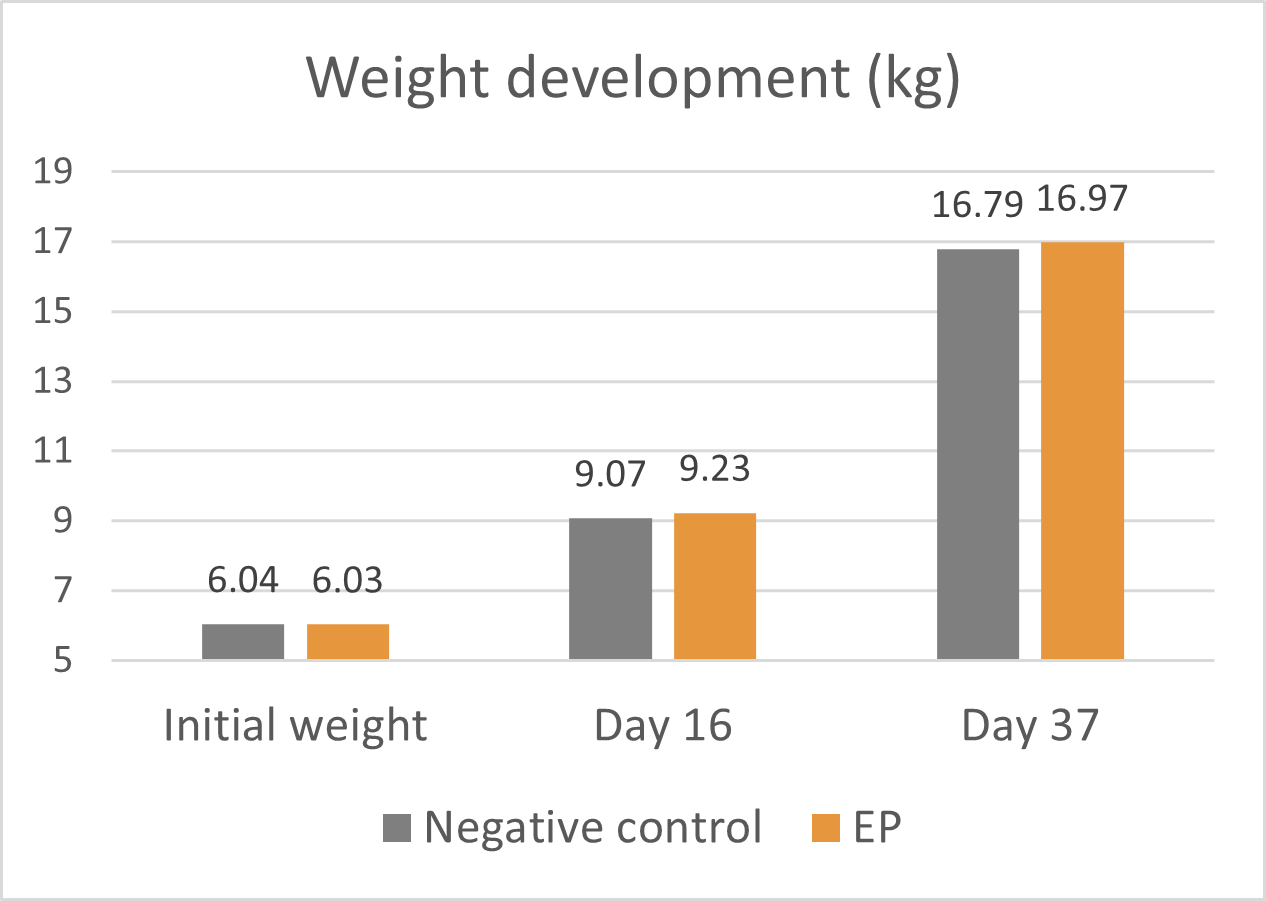
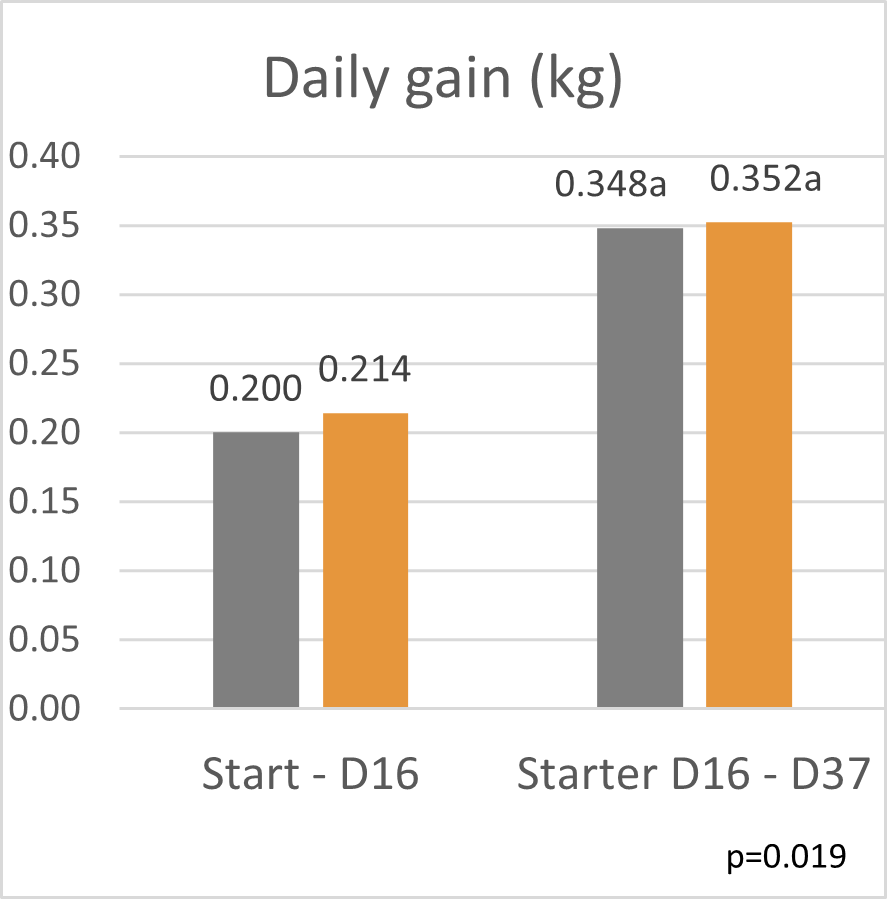
A trial conducted on a commercial farm in Spain shows the weight development of piglets fed an IgY-containing egg powder product (EP) compared to a negative control. The weaned piglets were fed a two-phase feeding (15 days prestarter, 22 days starter). The control (n=51) received no additional functional feed ingredient, whereas the EP group was fed 2 kg of the product/t of feed during the prestarter phase. The animals were weighed individually on days 16 and 37.
The results are shown in Figures 1 and 2.
Figure 1: Weight development of piglets receiving an IgY-containing egg powder product compared to a negative control
Figure 2: Daily gain of piglets receiving an IgY-containing egg powder product compared to a negative control
Explanation of the results: Under poor hygienic conditions, the pathogenic pressure is relatively high, and everything lowering this pressure helps to improve gut health, the utilization of nutrients, and performance. Egg immunoglobulins positively influence the gut microbiome, thus helping reduce diarrhea. By lowering the pathogenic pressure, the organism’s energy can be used for growth and must not be employed for the body’s defense.
7.2 Phytomolecules can even show improvement under optimum conditions
Phytomolecules generally show diverse gut health-promoting effects, from driving the intestinal microbiome in the right direction and strengthening the intestinal barrier to acting as antioxidants or anti-inflammatories or increasing the secretion of digestive juices and, therefore, improving digestion. Which mode of action is relevant if the piglets are raised under already optimal conditions (best hygiene, no prophylactic antibiotics or zinc oxide) and show the highest growth? Is there still room for improvement? Yes, it is. A trial conducted in Germany adduces evidence.
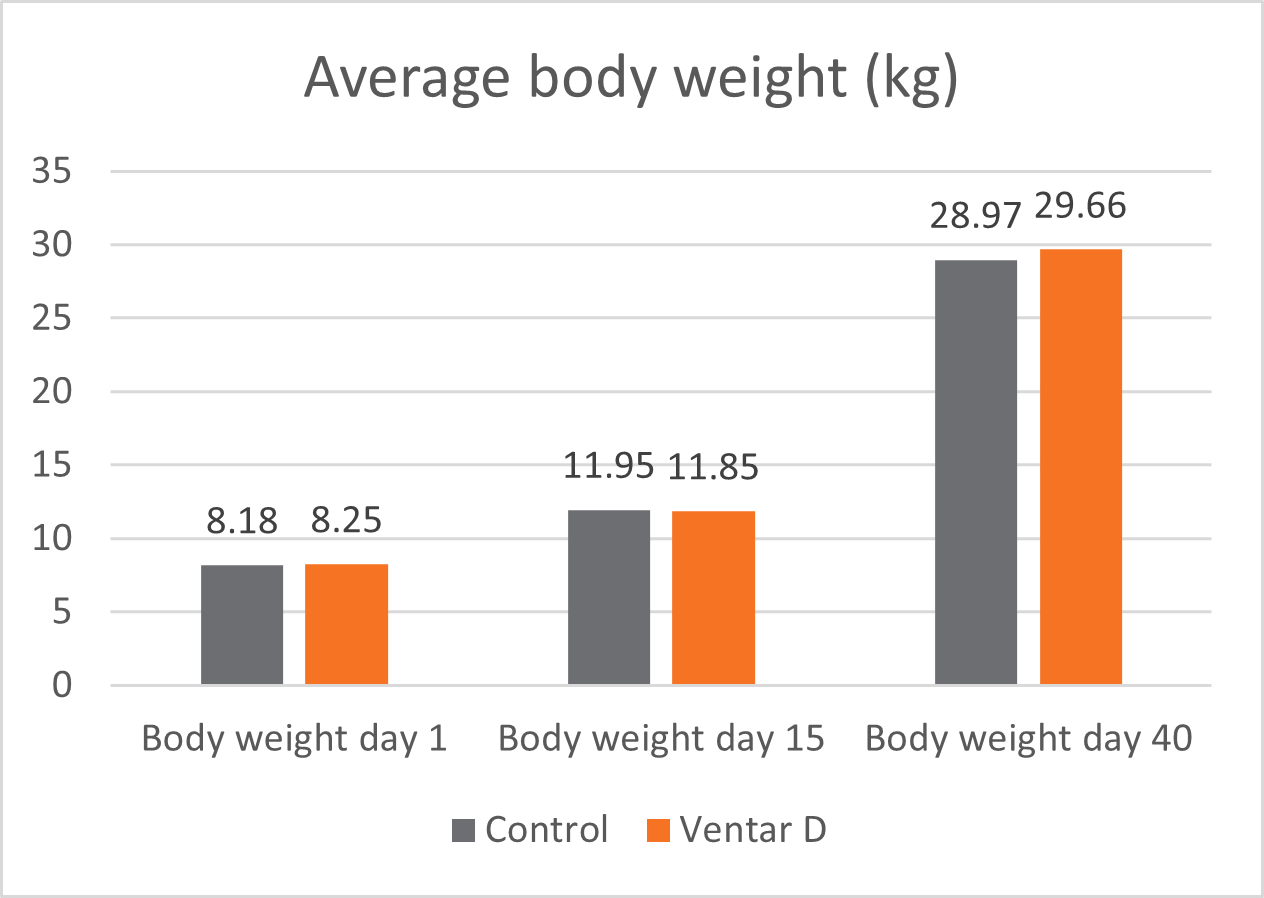
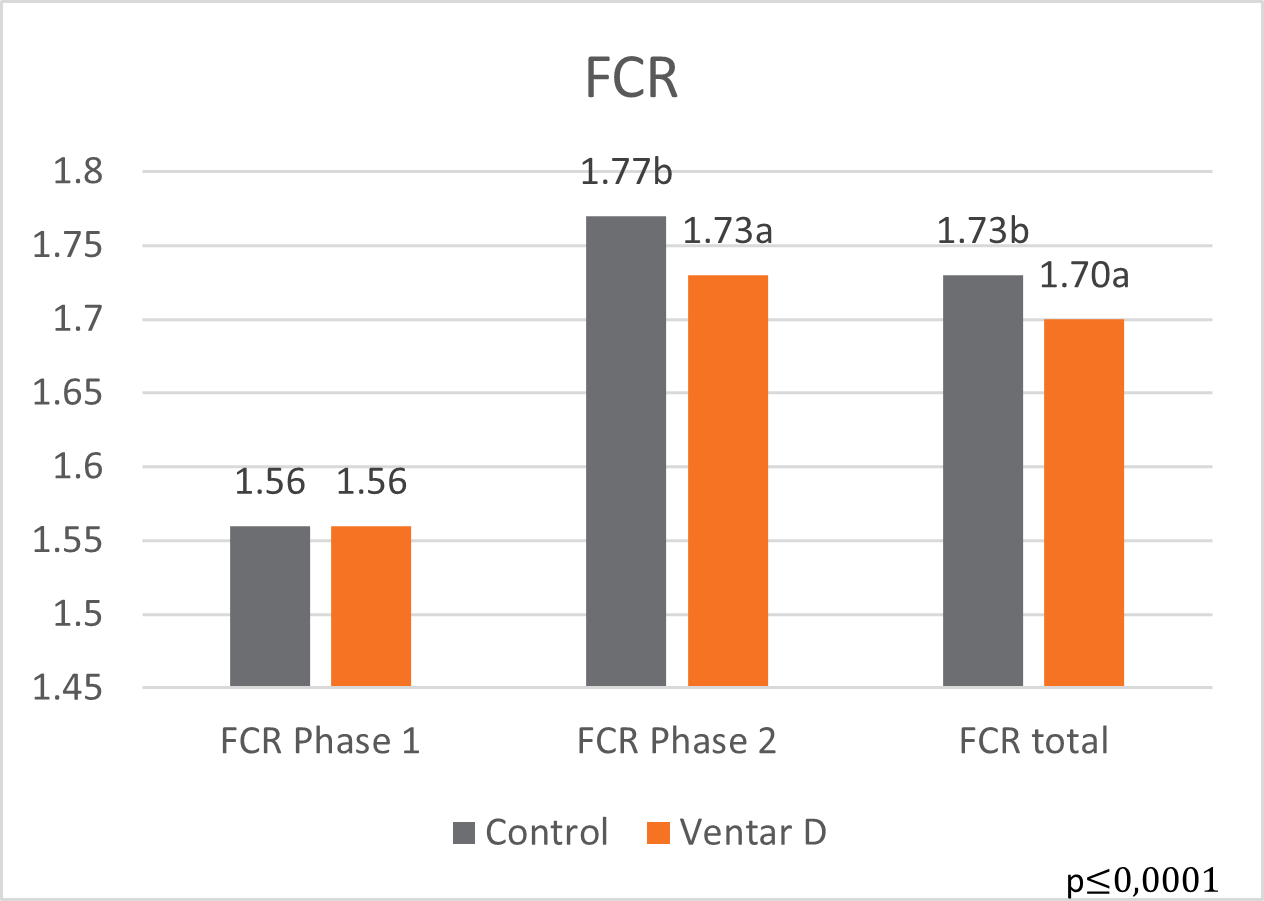
In this trial, 220 piglets weaned on average at 26 days and weighing around 8 kg were housed in 20 pens of 11 castrated males or gilts each. Piglets were blocked by body weight and fed a two-phase feeding program (phase 1 from day 1 to day 13 and phase 2 from day 17 to day 40; pelleted diet). Neither feed or water medication nor therapeutic levels of ZnO were used.
The results of this piglet trial can be seen in Figures 3 and 4.
Figure 3: Weight development of piglets fed Ventar D compared to a negative control
Figure 4: Feed conversion rate in piglets fed Ventar D compared to a negative control
Explanation of the results: The figures show that the piglets in the control already have an extremely high weight compared to those of a similar age in the previous trial, indicating the best rearing conditions in this trial. But, even here, Ventar D has the capacity to improve performance. Why? High-performing animals stress their body more than low-performing ones. Anabolic processes increase oxidative stress and non-infectious inflammation and burden the immune system. The relevant mode of action of Ventar D is not the gut health-promoting or the antimicrobial one because there is no issue. The relevant modes of action in this case are antioxidant and anti-inflammatory. With these two characteristics, Ventar D still has the capacity to improve the performance of piglets that are already at the top level.
8. Conclusion
For high piglet performance, providing the best possible rearing conditions is essential. However, there are differences concerning these conditions in different areas of the world. Depending on them, different feed strategies can be used. Egg immunoglobulins show the best effects if there is a certain pathogenic pressure. Phytomolecules, however, due to their various modes of action, can be beneficial under different levels in rearing conditions. In a low standard, the antimicrobial and gut health-promoting effect is more relevant; in the case of best conditions, the anti-oxidant and anti-inflammatory effects are decisive.
In summary, it could be said that functional feed ingredients have significant advantages in piglet rearing, but the right choice must be made depending on the prevailing conditions.
Sustainable livestock farming: Progress since 1950
by Ilinca Anghelescu, Global Director Marketing Communications, EW Nutrition
Summary
Global GHG Emissions and Agriculture:
Agriculture-related emissions account for 31% of global anthropogenic emissions, with a growing share coming from food-related activities outside traditional farming, such as processing and transportation.
This represents a significant decrease from the 1950s when agriculture contributed to 58% of global emissions, a decrease largely due to the increased use of fossil fuels.
Population Growth and Emissions:
The global population has increased by 220% since 1950, leading to a threefold increase in agri-food emissions, now totaling 9-10 billion metric tons of CO2-equivalent annually.
Meat Production Growth:
Meat production has seen a 690% increase since 1950, driven by population growth, economic development, urbanization, technological advancements, and intensification of livestock production.
Technological improvements have significantly increased livestock yield, including higher carcass weights, improved feed efficiency, and greater output per animal.
Feed Conversion Ratio (FCR) as a Sustainability Metric:
FCR, which measures the efficiency of feed conversion into body mass, has improved dramatically for poultry, pigs, and cattle since the 1950s.
Improved FCR contributes to more efficient resource use, reduced environmental impact, better animal welfare, and economic viability.
Livestock Emissions and Land Use:
Livestock-related emissions have increased by 14% since 2000, with a significant portion coming from enteric fermentation in ruminants and land use changes.
Pasture and grazing land have expanded slightly, while the land for feed crops has increased substantially due to intensified livestock production.
Food Loss and Its Impact on Sustainability:
Food loss, occurring primarily before the consumer stage, remains stable at around 13%. Reducing food loss is critical for improving food security, economic efficiency, and minimizing the environmental footprint of livestock production.
Future Strategies for Sustainability:
The article emphasizes the need for ongoing investment in technology, optimization of feed efficiency, sustainable land use, and improved methods for tracking and reducing emissions to ensure the future sustainability of livestock farming.
As the global demand for animal products continues to rise, so do various claims about the impact of agriculture on greenhouse gas emissions. A study commissioned by the United Nations’ Food and Agriculture Organization (FAO) concluded that, according to the most recent data, agri-food system emissions totaled 16.5 billion metric tons of CO2 equivalent, representing 31% of global anthropogenic emissions.
Of these 31%, the most important trend highlighted by FAO was the “increasingly important role of food-related emissions generated outside of agricultural land, in pre- and post-production processes along food supply chains”. The food supply chain (food processing, packaging, transport, household consumption and waste disposal) is thus set to become the top GHG emitter, above farming and land use.
How bad is 31%?
While 31% is a large figure, even this estimate represents a significant decrease from the 1950s, when agri-food emissions constituted approximately 58% of total anthropogenic emissions: “From 1850 until around 1950, anthropogenic CO2 emissions were mainly (>50%) from land use, land-use change and forestry”, states the latest IPCC report.
Figure 1. Source: IPCC AR6 Report, 2023. LULUCF = Land Use, Land-Use Change and Forestry
As the IPCC graph in Figure 1 indicates, the percentage decrease is mostly due to the rising prevalence of oil and coal in CO2 emissions over the recent decades, as shown in Figure 2 below.
Annual greenhouse gas (GHG) emissions worldwide from 1990 to 2022, by sector (in million metric tons of carbon dioxide equivalent)
Total population and agri-food emission changes, 1950 – today
The global population increased by approximately 220%, from 2.5 billion in 1950 to 8 billion in 2023. In the meantime, estimates suggest that, in the 1950s, agri-food systems were responsible for approximately 2-3 billion metric tons of CO2-equivalent (CO2e) emissions per year. This figure includes emissions from livestock, rice paddies, fertilizer use, and land-use change (e.g., deforestation for agriculture).
Assessments generally agree that today’s agri-food systems contribute approximately 9-10 billion metric tons of CO2e annually, a threefold increase from 1950. This includes emissions from agriculture (e.g., livestock, crop production), food processing, transportation, and land-use changes.
This increase is consistent with FAO’s new findings, of food chain climbing to the top of agri-food emitters.
But where did these increased emissions come from?
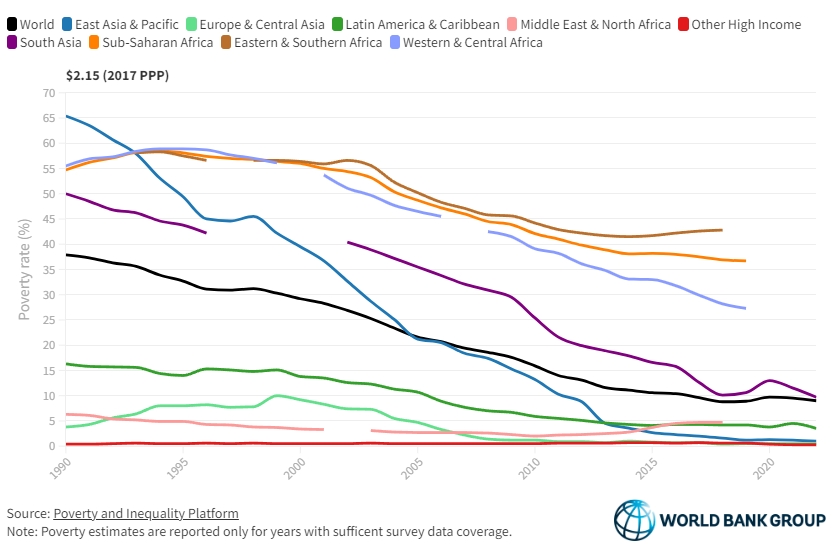
A look at the graph below gives us an indication: world poverty rate decreased massively between 1950 and today. While COVID brought a setback, the historical data would clearly indicate a correlation between the increased output in agri-food systems and the decreased rate of poverty.
How did poverty rates decline so steeply? The reasons lie, to a large extent, in technological innovation, especially in genetics and farm management, and in the increased apport of plentiful and affordable meat protein to the world. The numbers below build an image of an industry that produces better, more, and cheaper.
Global meat production: 1950 vs. Present
Then…
In 1950, the estimated total meat production was of approximately 45 million metric tons.
Key Producers: The United States, Europe, and the Soviet Union were the primary producers of meat. Types of Meat: Production was largely dominated by beef and pork, with poultry being less significant.
…and now
Now, the total meat production lies somewhere around 357 million metric tons (as of recent data from FAO)., representing a 53% increase from 2000 and a staggering 690% increase from 1950.
Key Producers: Major producers include China, the United States, Brazil, and the European Union. Types of Meat: Significant increases in poultry production, with pork remaining a leading source of meat, especially in Asia. Beef production has also increased, but at a slower rate than poultry and pork.
Factors contributing to increased meat production
Population Growth: The world population has grown from approximately 2.5 billion in 1950 to over 8 billion today, driving increased demand for meat.
Economic Growth and Urbanization: Rising incomes and urbanization have led to shifts in economic power and dietary preferences, with more people consuming higher quantities of meat, especially in developing countries.
Technological Advancements: Improvements in animal breeding, feed efficiency, and production systems have increased the efficiency and output of meat production.
Intensification of Livestock Production: The shift from extensive to intensive livestock production systems has allowed for higher meat yields per animal.
Global Trade: Expansion of global trade in meat and meat products has facilitated the growth of production in countries with comparative advantages in livestock farming.
Livestock yield increase, 1950 to the present
The increase in livestock yield for cattle, pigs, and chickens between 1950 and the present has been significant due to advances in breeding, nutrition, management practices, and technology.
Beef
1950s
Average Carcass Weight: In the 1950s, the average carcass weight of beef cattle was about 200 to 250 kilograms (440 to 550 pounds).
Dressing Percentage: The dressing percentage (the proportion of live weight that becomes carcass) was typically around 50-55%.
Present Day
Average Carcass Weight: Today, the average carcass weight of beef cattle is approximately 300 to 400 kilograms (660 to 880 pounds).
Dressing Percentage: The dressing percentage has improved to about 60-65%.
Increase in Beef Cattle Yield
Increase in Carcass Weight: The average carcass weight has increased by about 100 to 150 kilograms (220 to 330 pounds) per animal.
Improved Dressing Percentage: The dressing percentage has increased by about 5-10 percentage points, meaning a greater proportion of the live weight is converted into meat.
Dairy
1950s
Average Milk Yield per Cow: Approximately 2,000 to 3,000 liters per year, depending on the region.
Present Day
Average Milk Yield per Cow: Approximately 8,000 to 10,000 liters per year globally, with some countries like the United States achieving even higher averages of 10,000 to 12,000 liters per year.
Increase in Milk Yield:: Milk yield per cow has increased about 4-5 times due to genetic selection, improved nutrition, technological advancements, and better herd management.
Chickens (Layers)
1950s
Average Egg Production per Hen: In the 1950s, a typical laying hen produced about 150 to 200 eggs per year.
Present Day
Average Egg Production per Hen: Today, a typical laying hen produces approximately 280 to 320 eggs per year, with some high-performing breeds producing even more.
Increase in Egg Yield: The average egg production per hen has increased by approximately 130 to 170 eggs per year.
Chickens (Broilers)
1950s
Average Yield per Bird: In the 1950s, broiler chickens typically reached a market weight of about 1.5 to 2 kilograms (3.3 to 4.4 pounds) over a growth period of 10 to 12 weeks.
Present Day
Average Yield per Bird: Today, broiler chickens reach a market weight of about 2.5 to 3 kilograms (5.5 to 6.6 pounds) in just 5 to 7 weeks.
Increase in Yield: The average weight of a broiler chicken has increased by approximately 1 to 1.5 kilograms (2.2 to 3.3 pounds) per bird. Additionally, the time to reach market weight has been nearly halved.
Factors contributing to yield increases
Genetic Improvement:
Selective Breeding: Focused breeding programs have developed chicken strains with rapid growth rates and high feed efficiency, significantly increasing meat yield.
Nutrition:
Optimized Feed: Advances in poultry nutrition have led to feed formulations that promote faster growth and better health, using balanced diets rich in energy, protein, and essential nutrients.
Management Practices:
Housing and Environment: Improved housing conditions, including temperature and humidity control, have reduced stress and disease, enhancing growth rates.
Technological Advancements:
Automation: Automation in feeding, watering, and waste management has improved efficiency and bird health.
Health Monitoring: Advances in health monitoring and veterinary care have reduced mortality rates and supported faster growth.
Feed Conversion Efficiency:
Improved Feed Conversion Ratios (FCR): The amount of feed required to produce a unit of meat has decreased significantly, making production more efficient.
Why Feed Conversion Ratio is a sustainability metric
Feed Conversion Ratio (FCR) is a critical metric in livestock production that measures the efficiency with which animals convert feed into body mass. It is expressed as the amount of feed required to produce a unit of meat, milk, or eggs. Advances in nutrition and precision feeding allow producers to tailor diets that optimize FCR, reducing waste and improving nutrient uptake. Also, breeding programs focused on improving FCR can lead to livestock that naturally convert feed more efficiently, supporting long-term sustainability.
Poultry (Broilers): From the 1950s, improved from approximately 4.75 kg/kg to 1.7 kg/kg.
Pigs: From the 1950s, improved from about 4.5 kg/kg to 2.75 kg/kg.
Cattle (Beef): From the 1950s, improved from around 7.5 kg/kg to 6.0 kg/kg.
Figure 4. Evolution of FCR from 1950
FCR is crucial for livestock sustainability for several reasons, as shown below.
1. Resource efficiency
– Feed Costs: Feed is one of the largest operational costs in livestock production. A lower FCR means less feed is needed to produce the same amount of animal product, reducing costs and improving profitability.
– Land Use: Efficient feed conversion reduces the demand for land needed to grow feed crops, helping to preserve natural ecosystems and decrease deforestation pressures.
– Water Use: Producing less feed per unit of animal product reduces the water needed for crop irrigation, which is crucial in regions facing water scarcity.
2. Environmental impact
– Greenhouse Gas Emissions: Livestock production is a significant source of greenhouse gases (GHGs), particularly methane from ruminants and nitrous oxide from manure management. Improved FCR means fewer animals are needed to meet production goals, reducing total emissions.
– Nutrient Runoff: Efficient feed use minimizes excess nutrients that can lead to water pollution through runoff and eutrophication of aquatic ecosystems.
3. Animal welfare
– Health and Growth: Optimizing FCR often involves improving animal health and growth rates, which can lead to better welfare outcomes. Healthy animals grow more efficiently and are less susceptible to disease.
4. Economic viability
– Competitiveness: Lowering FCR improves the economic viability of livestock operations by reducing input costs and increasing competitiveness in the global market.
– Food Security: Efficient livestock systems contribute to food security by maximizing the output of animal protein relative to the input of resources.
Improving FCR is essential for achieving sustainability in livestock production. It leads to more efficient resource use, reduced environmental impact, enhanced economic viability, and supports the well-being of animals. As global demand for animal products continues to rise, optimizing FCR will be crucial in balancing production with the need to protect and preserve natural resources.
“The eight warmest years on record since 1961 (and in fact since the beginning of observations in 1880) are all within the eight-year period of 2015–2022. Europe is the region where the temperature change has been the highest in 2022 (and also for most of the 2000–2022 period), with 2.23 °C, followed by Asia (1.75 °C), the Americas (1.05 °C), Africa (1.01 °C) and Oceania (0.8 °C). The average temperature change in the 2010s was 1.25 °C, compared to 0.96 °C in the 2000s.” FAOSTAT 2023
Livestock emissions
Livestock emissions can be direct (farm-gate) or indirect (land use). Pre- and post-production emissions are considered separately, since they refer to emissions from manufacturing, processing, packaging, transport, retail, household consumption, and waste disposal.
Global farm-gate emissions (related to the production of crops and livestock) grew by 14% between 2000 and 2021, to 7.8 Gt CO2 eq, see below. 53% come from livestock-related activities, and the emissions from enteric fermentation generated in the digestive system of ruminant livestock were alone responsible for 37 percent of agricultural emissions (FAOSTAT 2023).
Land use emissions contribute a large share to agricultural emissions overall, especially through deforestation (~74% of land-use GHG emissions). The numbers have declined in recent years, to a total of 21% reduction between 2000 and 2018.
The other side of the coin is represented by the increased land usage for livestock, either directly for grazing or indirectly for feed crops.
1. Pasture and grazing land
1950: Approximately 3.2 billion hectares (7.9 billion acres) were used as permanent pastures.
Present: The area has increased to around 3.5 billion hectares (8.6 billion acres).
Change: An increase of about 0.3 billion hectares (0.7 billion acres).
2. Land for Feed Crops
1950: The land area dedicated to growing feed crops (such as corn and soy) was significantly less than today due to lower livestock production intensities and smaller scale operations. Feed crops likely accounted for about 200-250 million hectares of the cropland, although figures are evidently difficult to estimate.
Present: Of the approx. 5 billion hectares of land globally used for agriculture, about 1.5 billion hectares are dedicated to cropland.
The increase in cropland hectares is a direct consequence of the intensification of demand for livestock production. To keep these numbers in check, it is essential that producers strive to use as little feed as possible for as much meat yield as possible – and this directly relates to a key metric of the feed additive industry: Feed Conversion Ratio, mentioned above.
The role of food loss in livestock sustainability
The Food and Agriculture Organization (FAO) of the United Nations definesfood loss as the decrease in quantity or quality of food resulting from decisions and actions by food suppliers in the chain, excluding retail, food service providers, and consumers. Food loss specifically refers to food that gets spilled, spoiled, or lost before it reaches the consumer stage, primarily taking place during production, post-harvest, processing, and distribution stages.
Food loss is currently estimated to be relatively stable over the last decades, at around 13%.
Key aspects of food loss
Stages of Food Loss:
Production: Losses that occur during agricultural production, including damage by pests or diseases and inefficiencies in harvesting techniques.
Post-Harvest Handling and Storage: Losses that happen due to inadequate storage facilities, poor handling practices, and lack of proper cooling or processing facilities.
Processing: Losses during the processing stage, which may include inefficient processing techniques, contamination, or mechanical damage.
Distribution: Losses that occur during transportation and distribution due to poor infrastructure, inadequate packaging, and logistical inefficiencies.
Quality and Quantity:
Quality Loss: Refers to the reduction in the quality of food, affecting its nutritional value, taste, or safety, which may not necessarily reduce its quantity.
Quantity Loss: Refers to the actual reduction in the amount of food available for consumption due to physical losses.
Exclusions:
Retail and Consumer Level: Food loss does not include food waste at the retail or consumer levels, which is categorized as food waste. Food waste refers to the discarding of food that is still fit for consumption by retailers or consumers.
Importance of reducing food loss
Every step along the production chain, each action taken to preserve feed, increase yield, ensure stable and high meat quality, can contribute to reducing food loss and ensuring that animal protein production stays sustainable and feeds the world more efficiently.
Food Security: Reducing food loss can help improve food availability and access, particularly in regions where food scarcity is a concern. Where we thought we were on our way to eradicate world hunger, recent upticks in several regions show us that progress is not a given.
Economic Efficiency: Minimizing food loss can improve the efficiency and profitability of food supply chains by maximizing the utilization of resources.
Environmental Impact: Reducing food loss helps to decrease the environmental footprint of food production by lowering greenhouse gas emissions and minimizing land and water use. This is all the more important in regions where world hunger shows signs of going up. Perhaps not by coincidence are these regions some of the most affected by climate change.
By understanding and addressing the causes of food loss, stakeholders across the food supply chain can work towards more sustainable and efficient food systems.
What’s next?
Improving production practices and technology
Investment in research and development of new technologies that enhance livestock production efficiency and reduce environmental impact is vital for the future sustainability of the sector.
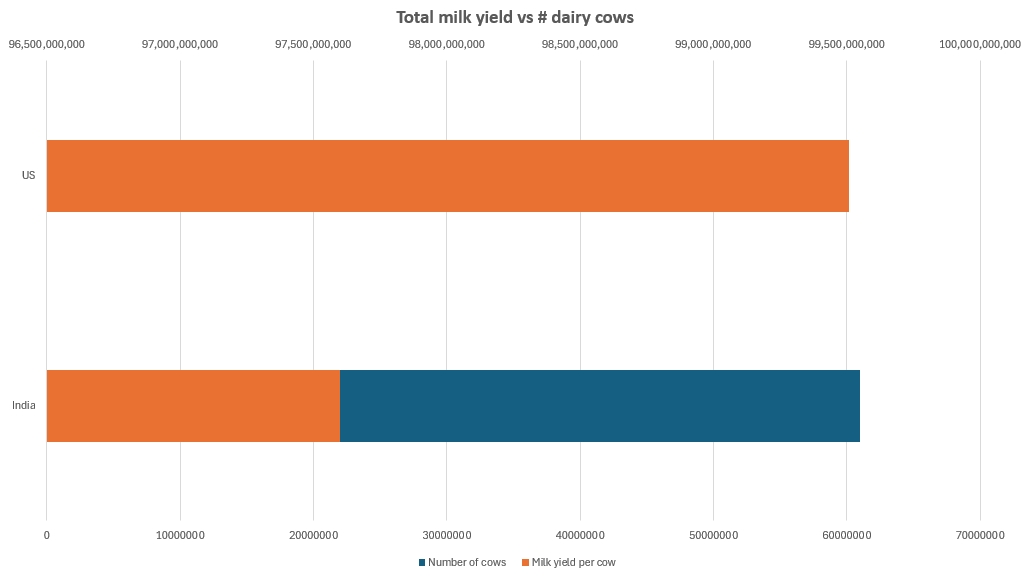
India is a good illustration of room to grow. If we look at cow milk alone, India, with a headcount of approximately 61 million animals, has a total milk production that is neck-and-neck with the United States, whose dairy cow headcount is in the neighborhood of 9.3 million. India’s milk yield sits around 1,600 liters/animal/year, compared to the US’s average of 10,700 liters.
Continued focus on improving FCR through genetic selection, optimized nutrition, and advanced management practices will be crucial for reducing the environmental footprint of livestock production.
Promoting Sustainable Land Use
Strategies to balance the need for increased livestock production with sustainable land use practices are essential. This includes adopting agroecological approaches and improving the efficiency of feed crop production.
Reducing Food Loss
Stakeholders across the food supply chain must prioritize reducing food loss through improved storage, transportation, and processing technologies. This will help ensure that livestock production contributes effectively to global food security.
Enhancing Emission Tracking and Reporting
There is a need for standardized methods for collecting and reporting data on GHG emissions in agriculture. This will enable more accurate assessments and the development of targeted strategies for emission reductions.
References
Bell, D. D. (2002). Laying hens in the U.S. market: An appraisal of trends from the beginning of the 20th century to present. Poultry Science, 81(5), 485-490. https://doi.org/10.1093/ps/81.5.485
Crippa, M., Solazzo, E., Guizzardi, D., Monforti-Ferrario, F., Tubiello, F. N., Leip, A., … & Janssens-Maenhout, G. (2022). Greenhouse gas emissions from food systems: building the global food system emissions database (GFED). Earth System Science Data, 14(4), 1795-1821. https://essd.copernicus.org/articles/14/1795/2022/essd-14-1795-2022.pdf
Goldewijk, K. K., & Verburg, P. H. (2013). Per-capita estimations of long-term historical land use and the consequences for global change research. Global Environmental Change, 23(4), 1166-1175. https://doi.org/10.1016/j.gloenvcha.2013.04.001
Kusuma, A. B., Laga, W. R., & Purnomo, H. (2022). Climate Change and Livestock Farming: Strategies for Mitigation and Adaptation. MDPI, 12(10), 1554. https://www.mdpi.com/2077-0472/12/10/1554
Roche, J. R., Friggens, N. C., Kay, J. K., Fisher, M. W., Stafford, K. J., & Berry, D. P. (2013). Invited review: Body condition score and its association with dairy cow productivity, health, and welfare. Animal Frontiers, 3(4), 23-29. https://doi.org/10.2527/af.2013-0032
Thornton, P. K., & Herrero, M. (2015). Impacts of climate change on the livestock food supply chain; a review of the evidence. Frontiers in Veterinary Science, 2, 93. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4686767/
Zuidhof, M. J., Schneider, B. L., Carney, V. L., Korver, D. R., & Robinson, F. E. (2014). Evolution of the modern broiler and feed efficiency. Annual Review of Animal Biosciences, 2(1), 47-71. https://doi.org/10.1146/annurev-animal-022513-114132
Global antimicrobial use in livestock farming: A revised estimate
Antimicrobial resistance (AMR) poses a significant threat to global health, driven by the overuse and misuse of antibiotics in both human medicine and livestock farming. In livestock farming, antimicrobials are still used extensively for therapeutic and non-therapeutic purposes. However, estimates of the quantities used per species are notoriously hard to derive from fragmented, incomplete, or unstandardized data around the world.
A recent article (“Global antimicrobial use in livestock farming: an estimate for cattle, chickens, and pigs”, Animal, 18(2), 2024) attempts to update the figures by estimating global biomass at treatment of cattle, pigs, and chickens, considering distinct weight categories for each species in biomass calculation, and using the European Medicines Agency’s weight standards for the animal categories. With these more refined calculations, authors Zahra Ardakani, Maurizio Aragrande, and Massino Canali aim to provide a more accurate estimate of global antimicrobial use (AMU) in cattle, chickens, and pigs. Understanding these patterns is crucial for addressing AMR and developing strategies for sustainable livestock management.
Key Findings
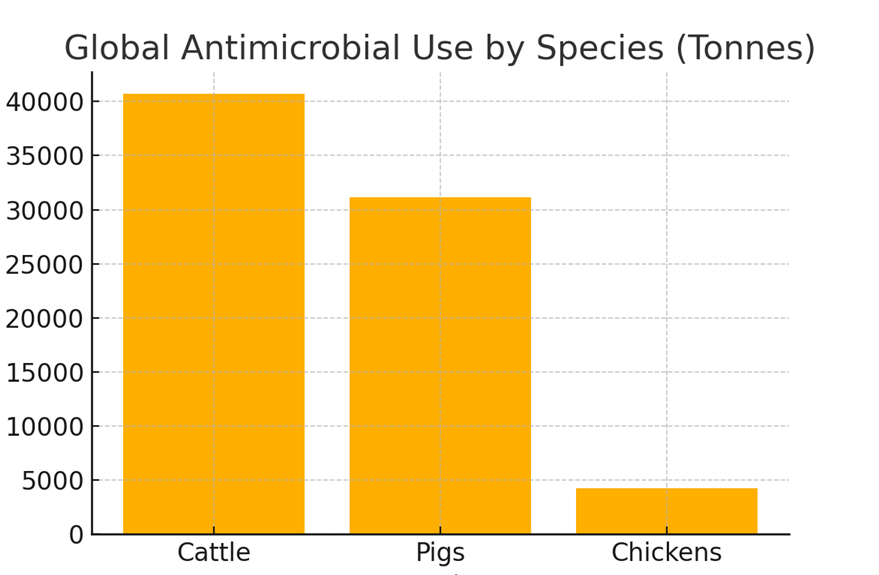
The study estimates that the global annual AMU for cattle, chickens, and pigs amounts to 76,060 tons of antimicrobial active ingredients. This is a significant revision from previous estimates due to a more detailed evaluation of animal weights and categories:
1. Cattle: 40,697 tons (53.5% of total AMU)
2. Pigs: 31,120 tons (40.9% of total AMU)
3. Chickens: 4,243 tons (5.6% of total AMU)
Figure 1: Distribution of global antimicrobial use among cattle, pigs, and chickens.
Methodology
The study utilizes the concept of Population Correction Units (PCU) to estimate antimicrobial usage, taking into account the weight and category of livestock at the time of treatment. This method differs from previous approaches that relied on live weight at slaughter, providing a more accurate representation of AMU.
The PCU is calculated by multiplying the number of animals by their average weight during treatment. This approach allows for differentiation by age and sex, which is particularly important for species like cattle and pigs.
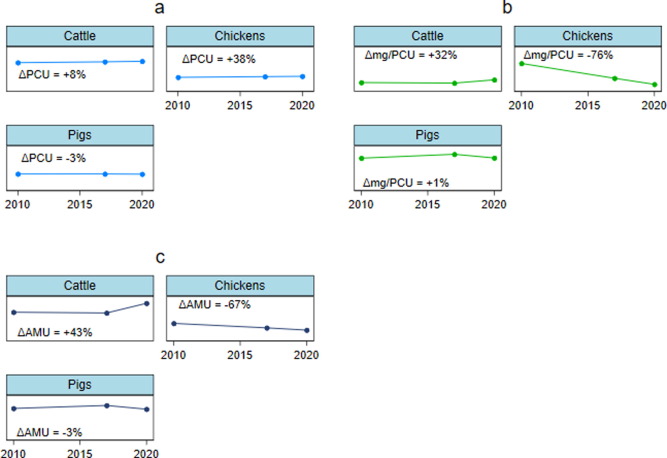
Figure 2: (a) Changes in global PCU (million tonnes), (b) changes in global antibiotic use in mg per PCU, and (c) changes in global AMU (thousand tonnes) for cattle, chickens, and pigs; between 2010 and 2020. Abbreviations: PCU = Population Correction Unit; AMU = Antibiotic Use.
Figure 2: (a) Changes in global PCU (million tonnes), (b) changes in global antibiotic use in mg per PCU, and (c) changes in global AMU (thousand tonnes) for cattle, chickens, and pigs; between 2010 and 2020. Abbreviations: PCU = Population Correction Unit; AMU = Antibiotic Use.
Study shows lower AMU than previous estimates
The study highlights a significant shift in AMU patterns, with chickens showing a remarkable decrease in antimicrobial use despite increased production. This is indicative of improved management and more responsible use of antibiotics in the poultry industry.
The lower AMU in cattle and pigs, compared to previous estimates, underscores the importance of considering animal age and weight at treatment. These findings align closely with World Organization for Animal Health (WOAH) estimates, validating the methodology.
However, the study also acknowledges limitations, including reliance on European standards for average weight at treatment, which may not reflect global variations. Additionally, the lack of comprehensive global data on veterinary antibiotics presents challenges in creating fully accurate estimates.
Corrected estimate highlights improved production advances
This study provides a revised and potentially more accurate estimate of global antimicrobial use in livestock. By accounting for the weight and treatment categories of animals, it offers insights that could guide policy and management practices to mitigate the spread of antimicrobial resistance.
The article also indicates that the industry may have over-estimated antimicrobial usage in livestock and, just as importantly, that antimicrobial use has been kept in check or even reduced, despite increases in farmed animal headcounts. The lower usage is likely due to regulatory oversight and improvements in alternative methods to control and mitigate health challenges.
The Global and European Pork Industry: An Overview from IFIP
Institut du Porc (IFIP), the French pork research and development institute and a key player in the French agricultural sector, has recently published its 2024 report, titled Porc par les Chiffres 2023-2024. The document provides a comprehensive overview of the global, European, and French pork industries and is a critical resource for all industry professionals.
The Global Pork Industry
The global pork industry has experienced significant changes over recent years. In 2021, global pork production reached approximately 108 million tons of carcass weight equivalent (CWE), largely driven by China’s recovery from African Swine Fever (ASF). By 2022, global production continued to rise, though at a slower pace due to the stabilization of China’s pig herd. However, the global landscape remains varied with notable regional differences.
In Asia, China dominates pork production, contributing 47.5 million tons in 2021. Other significant producers include Vietnam, Japan, South Korea, and the Philippines. In Europe, the EU-28 collectively produced 24.6 million tons, with Germany, Spain, and France being the leading producers. In the Americas, the United States and Brazil are major contributors, with the U.S. producing 12.6 million tons and Brazil 4.4 million tons. The production in other regions like Africa and Oceania remains relatively small in comparison.
The global pork trade is equally dynamic. In 2022, the major exporters included the European Union, the United States, and Canada. Key importers were China, Japan, and Mexico. The shifting demands and production capacities have led to fluctuating trade patterns, impacting global pork prices and market stability.
Figure 3. Evolution of global pork production (GDP in 1000 tons)
Pork Industry in the European Union
The European pork industry faced a challenging year in 2022, marked by a decline in production and various economic pressures. The EU’s pork production fell by approximately 5%, equating to a loss of over 12.6 million pigs for slaughter. Germany, historically one of the largest producers, saw a significant 10% reduction in output due to ongoing economic and sanitary crises. Spain, while typically a growing market, experienced its first production decline since 2014 due to increased piglet mortality rates from health issues.
Despite these challenges, some regions showed resilience. France managed a relatively smaller production decrease of 2%, maintaining its position as a key player in the European market. The economic environment, characterized by rising inflation and high feed costs, pushed pork prices to record levels across the continent, with French pork prices ranking high in Europe, just behind Spain.
The consumption patterns within the EU also varied significantly. Countries like Denmark and Spain exhibited high per capita pork consumption rates, while others like the United Kingdom and Italy showed more moderate consumption levels. This disparity reflects both cultural preferences and economic conditions across the region.
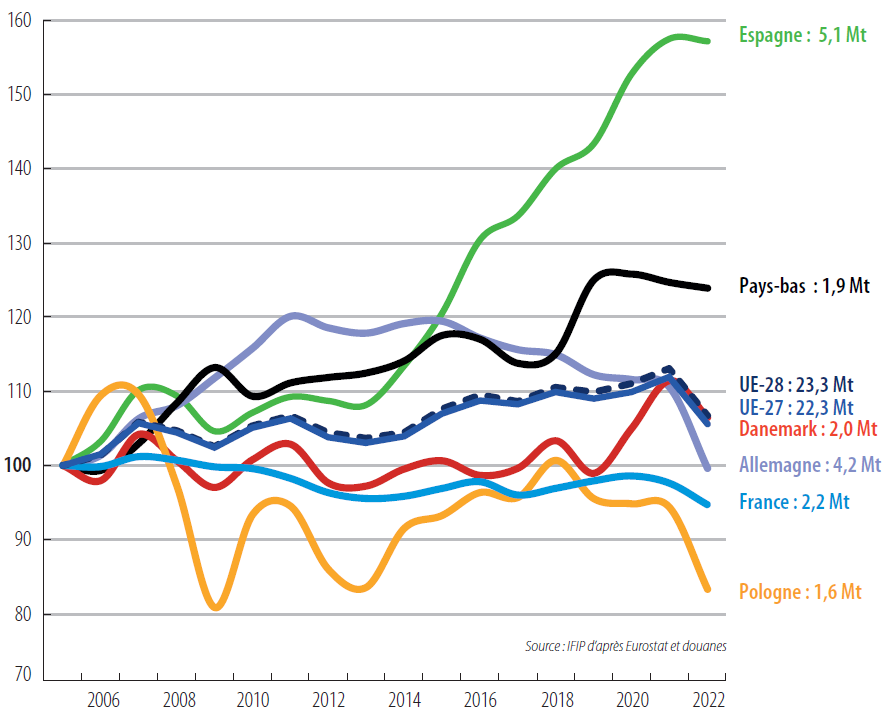
Figure 4. Evolution of pork production in the main EU countries (tons, from a base of 100 in 2005)
The French Pork Industry
In France, the pork industry in 2022 faced a year of significant adjustments. The country produced 2.19 million tons of carcass weight equivalent, a 3% decrease from the previous year. This decline was attributed to reduced slaughter weights and lower export volumes of live pigs. Despite these challenges, France remained the third-largest pork producer in the EU, following Spain and Germany.
Regionally, pork production is concentrated in areas like Brittany and Pays de la Loire. Brittany alone accounts for a substantial portion of the national production. The distribution of pork farms across France highlights the regional specialization, with significant variations in production volumes from one region to another.
Figure 5. Suppliers to France (percentage of total imports)
French pork exports faced hurdles due to reduced demand from China, which saw a 35% drop in imports from France in 2022. However, increased sales to other Asian markets like the Philippines and Japan partially offset this decline. In terms of value, the higher pork prices helped mitigate the impact of lower export volumes, with total export values reaching 1.76 billion euros.
Figure 6. Export targets for France (percentage of total exports)
Economic and Production Challenges
The pork industry globally and within the EU faces several ongoing challenges. Rising feed costs, largely driven by global commodity price increases, have significantly impacted production costs. In Europe, the economic downturn and ongoing health crises like ASF and PRRS (Porcine Reproductive and Respiratory Syndrome) continue to challenge producers.
In France, inflation and high production costs have led to a tightening of profit margins for pig farmers. The high costs of feed and energy, coupled with lower production volumes, have made it difficult for many producers to remain profitable. The industry has responded with efforts to improve efficiency and sustainability, though these measures take time to implement and yield results.
Future Outlook
In Asia, China’s recovery from ASF will likely stabilize, but the focus will shift towards improving biosecurity and production efficiency. In Europe, the industry will need to navigate economic challenges and health crises while adapting to changing consumer preferences towards more sustainable and ethical production practices.
For France, the key to future success will lie in balancing production efficiency with market demands. Investments in technology, biosecurity, and sustainable practices will be crucial. Additionally, expanding export markets beyond traditional partners will help mitigate the risks associated with market fluctuations.
The pork industry, both globally and within the EU, is at a pivotal point. The combination of economic pressures, health challenges, and shifting market dynamics necessitates strategic adjustments. By focusing on efficiency, sustainability, and market diversification, the industry can navigate these challenges and continue to thrive in the coming years.
Oxidative & Inflammatory stress in reproductive Sows
By Twan van Gerwe, DVM PhD Dipl ECPVS, Technical Director Dr. Inge Heinzl, Editor
One of the biggest challenges in swine production is keeping the modern, hyperprolific sow healthy and in good shape so that she can wean large, healthy litters and maintain her high reproductive performance.
Unfortunately, sows often suffer from stress and increased systemic inflammation around farrowing and during lactation. This leads to impaired feed intake and disturbed endocrine homeostasis, negatively affecting reproductive and litter performance.
The key to increasing the efficiency of pig production is to reduce the metabolic burden of sows while maintaining the reproductive performance of high-yield sows. A deep understanding of the complex interplay between environmental factors, sow well-being, health, and productivity is necessary to implement enhanced nutritional regimens and meticulous management practices.
Why does oxidative stress occur in today’s sows?
Nowadays, hyperprolific sows produce between 30 and 40 weaned piglets per year and are at a higher risk of suffering from stress. What are the reasons?
A high number of piglets causes oxidative stress
Oxidative stress occurs when reactive oxygen species (ROS) are produced faster than the body’s antioxidant mechanisms can neutralize them and cause damage to lipids, proteins, and DNA. During gestation, the sow needs high amounts of energy to provide for the fetuses. This energy is produced in the placental mitochondria. The placenta, therefore, is a place of active oxygen metabolism during gestation and a source of oxidative stress. In hyperprolific sows, a higher number of fetuses need even more energy to grow. Consequently, ROS production and the risk for intrauterine growth retardation (IUGR) increases (Figure 1). Moreover, evidence shows that the body’s antioxidant potential is reduced in late gestation and after parturition (Szczubial, 2010), resulting in increased oxidative stress biomarkers (Yang, 2023). Increased milk production for large litters demands a substantial amount of energy, risking similar oxidative distress. Therefore, both the final phase of gestation and the subsequent lactation period are predestined for oxidative stress, which has been demonstrated by reduced TEAC (Trolox equivalent antioxidant capacity) levels during these phases (Lee et al., 2023).
Figure 1. Illustration of the effect of oxidative stress on the fetus: intrauterine growth retardation (IUGR) (adapted from Yang et al., 2023)
Heat and ambient stress also contribute
The reproductive sow produces lots of heat. From the beginning of gestation, the sow’s thermoneutral zone decreases. This, however, does not always correspond with the ambient conditions. Especially during the last days of gestation, the discrepancy is exceptionally high as everything is prepared for the newborn piglets, which need a temperature of about 27-35°C. The sow, on the contrary, would be happy with 18-22°C. Additionally, changes around farrowing – moving to the farrowing unit, social stress, change of feed, and the preparation for parturition – exert additional stress for the sows.
Why does the inflammation level increase?
After parturition, systemic inflammation is a normal phenomenon: the reproductive organs have sustained injuries during the parturition process and require remodeling. Inflammation is a natural and desired process, to repair the tissues and return to a normal status. However, inflammation is increased in modern sows, adversely affecting their inflammatory balance. Some possible underlying reasons are:
The high numbers of piglets need a lot of space in the uterus, often leading to damage of the uterine tissue and an inflammatory response in the sows. Lee et al. (2023) found significantly (p<0.10) higher TNF-α concentrations in sows with litters of 15-20 piglets than in sows with 7-14 piglets. TNF-α is a biomarker of inflammation.
Pathogenic infections – particularly infections of the reproductive tract – can induce a prolonged or excessive inflammatory state. A further reason can be the need for more obstetric interventions in hyperprolific sows, which can injure the birth canal or the uterus.
Imbalanced nutrition: Excessive backfat is associated with a higher expression of proinflammatory cytokines, and feed contaminated with mycotoxins can impair the sow’s immunocompetence.
Biomarkers can inform us about the oxidative status
Biomarkers are naturally occurring molecules that help us identify diseases or physiological processes. They provide insights into the oxidative state and inflammatory processes.
Anti-oxidative biomarkers
To check the anti-oxidative capacity, the “beneficial” substances, or antioxidants, can be quantified. These substances can neutralize free radicals or be neutralized by them. Higher levels of antioxidants indicate better antioxidant capacity; when antioxidants are abundant, fewer oxidizable substances have undergone oxidation.
Examples of antioxidant biomarkers:
Total Antioxidant Capacity (T-AOC): represents the synergistic interaction effects of all antioxidants in a matrix (E.g., diet or body fluids). It’s a global measure of non-enzymatic antioxidant efficiency. Various assays, like Trolox Equivalent Antioxidant Capacity (TEAC), which measures a substance’s antioxidant capacity compared to Trolox, can measure T-AOC.
Glutathione Peroxidase (GSH-Px) belongs to the peroxidase family and converts hydrogen peroxide to water.
Catalase (CAT): scavenges ROS. Its activity can predict oxidative stress.
Superoxide Dismutase (SOD): catalyzes the dismutation of superoxide radicals to oxygen and hydrogen peroxide.
Oxidative biomarkers
Oxidative stress biomarkers, the ‘negative’ substances, can also serve as general biomarkers. These include free radicals with oxidant capacity or intermediate/final oxidation products. Ideally, their levels should be minimized.
Examples of oxidative stress biomarkers:
Thiobarbituric acid reactive substances (TBARS): to measure lipid peroxidation products in cells, tissues, and body fluids.
Reactive oxygen species(ROS) or free radicals: unstable, oxygen-containing molecules that react with other molecules in a cell. They might damage DNA, RNA, and proteins and cause cell death. Hydrogen Peroxide (H₂O₂) is a ROS produced during normal cellular metabolism, which causes oxidative damage at excessive levels.
Malondialdehyde (MDA): a final product of oxidative fat degradation and, therefore, a biomarker for lipid peroxidation.
Pro-inflammatory biomarkers
Like oxidative stress, the interplay between pro- and anti-inflammatory signals helps develop the proper immune response for the appropriate duration.
Examples of Pro-inflammatory biomarkers or molecules produced in the case of inflammation:
Plasma Adenosine Deaminase (ADA-1 and ADA-2): involved in immune regulation, with ADA-1 inhibiting pro-inflammatory responses and ADA-2 supporting immune cell functions.
Interleukins (IL-1α and IL-1β), IL-6: IL-1α and IL-1β are associated with inflammatory diseases, IL-6: is produced during inflammation and acute-phase response.
Tumor Necrosis Factor α (TNF-α): endogenous pyrogen that induces fever and promotes inflammation.
C-reactive Protein (CRP): liver-produced acute-phase protein responding to inflammation.
Procalcitonin (PCT) is produced by the liver during infections and helps detect bacterial infections.
Examples of anti-inflammatory substances – the “good ones”:
Interleukines – IL-4, IL-10: inhibit the function of the macrophages and act, therefore, anti-inflammatory
Cortisol: anti-inflammatory and immune-suppressive
ACTH: stimulates the production and release of cortisol
Higher stress or infection level lowers performance in sows and piglets
As mentioned, hyperprolific sows suffer from higher oxidative stress, especially during late gestation, parturition, and lactation. Additionally, systemic inflammation occurs to repair the injured tissues to facilitate the healing of the birth canal and remodeling of the uterus to establish the subsequent pregnancy. To this purpose, an inflammatory cascade, triggered by the injuries due to gestation and parturition, involves the release of critical (pro-inflammatory) mediators such as TNF-α and IL-6, leading to the activation of acute phase proteins.
After triggering inflammatory pathways, anti-inflammatory pathways must also be activated to reestablish homeostasis in the reproductive organs (Serhan & Chiang, 2008). Alterations at the onset of anti-inflammatory pathways and exacerbated activation and maintenance of inflammatory pathways can lead to uncontrolled inflammation and the onset of reproductive disease in sows (Kaiser et al., 2018), as well as reduced feed intake and insufficient milk production, resulting in poorly growing piglets and lower weaning weights or piglets suffering from clinical infectious diseases such as diarrhea. If possibly homeostasis cannot be restored, the sow is at risk of contracting diseases like post-partum dysgalactia syndrome (PPDS), lameness, and impaired fertility.
Targeted use of polyphenols can mitigate inflammation and improve the oxidative status of sows
There are several experiments showing the beneficial effects of natural compounds. Especially polyphenols, disposing of phenyl rings and two or more hydroxyl substituents, are perfect radical scavengers and proven antioxidants (Chen, 2023). Phytogenic substances that have anti-inflammatory effects can be found in the families of polyphenols as well as terpenoids, flavonoids, saponins, and tannins (Bunte et al., 2019; Ge et al., 2022; Ginwala et al., 2019; Santos Passos et al., 2022; Ambreen and Mirza, 2020).
Here are some examples showing the beneficial effects of phytochemicals:
Primiparous sows fed with Moringa oleifera leaf meal, rich in polyphenols, saponins, and tannins, illustrate the potential of phytomolecules: serum levels of T-AOC (total anti-oxidative capacity), were increased in late gestation and during lactation, while MDA was reduced. Additionally, piglets that received Moringa oleifera meal showed the highest serum CAT and SOD activities. The meal significantly decreased the farrowing length and number of stillbirths, while there was an increasing trend in the number of live‐born piglets (Sun et al., 2020).
The polyphenol Daidzein, a member of the class of compounds known as isoflavones (200 mg/kg during gestation), increased the total antioxidant capacity (T-AOC) and the activities of glutathione peroxidase and superoxide dismutase. Additionally, it elevated the level of immunoglobulin G and increased the number of piglets born and born alive per litter (Li et al., 2021).
Glycitein, a polyphenol occurring in the isoflavone fraction of soy products, applied during late gestation and lactation increased the total antioxidant capacity and SOD activity during the first 18 days of lactation and the CAT and GSH-Px activity in mid-lactation. Plasma MDA level was reduced from late gestation to the 18th day of lactation. The enhanced oxidative status of the sow resulted in a higher daily gain of the piglets and a higher weaning weight of the litter (Hu et al., 2015).
Meng et al. (2018) tested Resveratrol (300 mg/kg), a stilbenes polyphenol, in sows from day 20 of gestation until farrowing. They saw noticeably higher GSH-Px, SOD, and CAT activities, as well as lower contents of MDA and H2O2 in the placental tissue, improving the antioxidant status of sows and piglets.
Xu et al. (2022) fed silymarin to sows in late gestation. They observed that IL-1ß concentration in the blood sample on the 18th day of lactation was reduced in the supplemented group. The altered fecal microbiota was associated with variations in inflammatory factors, suggesting that silymarin modulates microbiota in the gut and may improve the health of lactation sow.
Phytochemicals support sows against oxidative and inflammatory stress
The above-presented examples show that phytochemicals, particularly those developed to have a potent anti-inflammatory and anti-oxidative capacity, have a high potential to alleviate oxidative stress in pregnant and lactating sows and reduce inflammation when applied in sow diets. Consequently, a broader use of these natural substances should be considered to reduce the metabolic burden of sows and increase the efficiency of pig production.
References:
Ambreen, Madieha, and Safdar Ali Mirza. “Evaluation of Anti-Inflammatory and Wound Healing Potential of Tannins Isolated from Leaf Callus Cultures of Achyranthes Aspera and Ocimum Basilicum.” Pak J Pharm Sci . 33, no. 1 (January 2020): 361–69.
Bunte, Kübra, Andreas Hensel, and Thomas Beikler. “Polyphenols in the Prevention and Treatment of Periodontal Disease: A Systematic Review of in Vivo, Ex Vivo and in Vitro Studies.” Fitoterapia 132 (January 2019): 30–39. https://doi.org/10.1016/j.fitote.2018.11.012.
Chen, Jun, Zhouyin Huang, Xuehai Cao, Tiande Zou, Jinming You, and Wutai Guan. “Plant-Derived Polyphenols in Sow Nutrition: An Update.” Animal Nutrition 12 (March 2023): 96–107. https://doi.org/10.1016/j.aninu.2022.08.015.
Ge, Jiamin, Zhen Liu, Zhichao Zhong, Liwei Wang, Xiaotao Zhuo, Junjie Li, Xiaoying Jiang, Xiang-Yang Ye, Tian Xie, and Renren Bai. “Natural Terpenoids with Anti-Inflammatory Activities: Potential Leads for Anti-Inflammatory Drug Discovery.” Bioorganic Chemistry 124 (July 2022): 105817. https://doi.org/10.1016/j.bioorg.2022.105817.
Ginwala, Rashida, Raina Bhavsar, De Gaulle Chigbu, Pooja Jain, and Zafar K. Khan. “Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin.” Antioxidants 8, no. 2 (February 5, 2019): 35. https://doi.org/10.3390/antiox8020035.
Hu, Y. J., K. G. Gao, C. T. Zheng, Z. J. Wu, X. F. Yang, L. Wang, X. Y. Ma, A. G. Zhou, and Z. J. Jiang. “Effect of Dietary Supplementation with Glycitein during Late Pregnancy and Lactation on Antioxidative Indices and Performance of Primiparous Sows1.” Journal of Animal Science 93, no. 5 (May 1, 2015): 2246–54. https://doi.org/10.2527/jas.2014-7767.
Kaiser, Marianne, Stine Jacobsen, Pia Haubro Andersen, Poul Bækbo, José Joaquin Cerón, Jan Dahl, Damián Escribano, Peter Kappel Theil, and Magdalena Jacobson. “Hormonal and Metabolic Indicators before and after Farrowing in Sows affected with postpartum Dysgalactia Syndrome.” BMC Veterinary Research 14, no. 1 (November 7, 2018). https://doi.org/10.1186/s12917-018-1649-z.
Lee, Juho, Hyeonwook Shin, Janghee Jo, Geonil Lee, and Jinhyeon Yun. “Large Litter Size Increases Oxidative Stress and Adversely Affects Nest-Building Behavior and Litter Characteristics in Primiparous Sows.” Frontiers in Veterinary Science 10 (August 22, 2023). https://doi.org/10.3389/fvets.2023.1219572.
Li, Yan, Guoru He, Daiwen Chen, Bing Yu, Jie Yu, Ping Zheng, Zhiqing Huang, et al. “Supplementing Daidzein in Diets Improves the Reproductive Performance, Endocrine Hormones and Antioxidant Capacity of Multiparous Sows.” Animal Nutrition 7, no. 4 (December 2021): 1052–60. https://doi.org/10.1016/j.aninu.2021.09.002.
Meng, Qingwei, Tao Guo, Gaoqiang Li, Shishuai Sun, Shiqi He, Baojing Cheng, Baoming Shi, and Anshan Shan. “Dietary Resveratrol Improves Antioxidant Status of Sows and Piglets and Regulates Antioxidant Gene Expression in Placenta by Keap1-Nrf2 Pathway and SIRT1.” Journal of Animal Science and Biotechnology 9, no. 1 (April 20, 2018). https://doi.org/10.1186/s40104-018-0248-y.
Santos Passos, Fabiolla Rocha, Heitor Gomes Araújo-Filho, Brenda Souza Monteiro, Saravanan Shanmugam, Adriano Antunes Araújo, Jackson Roberto Almeida, Parimelazhagan Thangaraj, Lucindo José Júnior, and Jullyana de Quintans. “Anti-Inflammatory and Modulatory Effects of Steroidal Saponins and Sapogenins on Cytokines: A Review of Pre-Clinical Research.” Phytomedicine 96 (February 2022): 153842. https://doi.org/10.1016/j.phymed.2021.153842.
Serhan, C N, and N Chiang. “Endogenous Pro‐resolving and Anti‐inflammatory Lipid Mediators: A New Pharmacologic Genus.” British Journal of Pharmacology 153, no. S1 (March 2008). https://doi.org/10.1038/sj.bjp.0707489.
Sun, Jia‐Jie, Peng Wang, Guo‐Ping Chen, Jun‐Yi Luo, Qian‐Yun Xi, Geng‐Yuan Cai, Jia‐Han Wu, et al. “Effect of Moringa Oleifera Supplementation on Productive Performance, Colostrum Composition and Serum Biochemical Indexes of Sow.” Journal of Animal Physiology and Animal Nutrition 104, no. 1 (October 30, 2019): 291–99. https://doi.org/10.1111/jpn.13224.
Szczubiał, M. “Changes in Oxidative Stress Markers in Plasma of Sows during Periparturient Period.” Polish Journal of Veterinary Sciences, March 3, 2020, 185–90. https://doi.org/10.24425/pjvs.2020.132764.
Xu, Shengyu, Xiaojun Jiang, Xinlin Jia, Xuemei Jiang, Lianqiang Che, Yan Lin, Yong Zhuo, et al. “Silymarin Modulates Microbiota in the Gut to Improve the Health of Sow from Late Gestation to Lactation.” Animals 12, no. 17 (August 26, 2022): 2202. https://doi.org/10.3390/ani12172202.
Yang, Xizi, Ruizhi Hu, Mingkun Shi, Long Wang, Jiahao Yan, Jiatai Gong, Qianjin Zhang, Jianhua He, and Shusong Wu. “Placental Malfunction, Fetal Survival and Development Caused by Sow Metabolic Disorder: The Impact of Maternal Oxidative Stress.” Antioxidants 12, no. 2 (February 2, 2023): 360. https://doi.org/10.3390/antiox12020360.
Unlocking Optimum Poultry Performance: Harnessing the Power of GH10 Xylanase
Author: Ajay Bhoyar, Global Technical Manager, EW Nutrition
Exogenous feed enzymes are increasingly utilized in poultry diets to manage feed costs, mitigate the adverse effects of anti-nutritional factors, and enhance nutrient digestion and bird performance. These enzymes are primarily employed to bolster the availability of nutrients within feed ingredients. Among the various enzymes utilized, those capable of breaking down crude fiber, starch, proteins, and phytates are commonly integrated into animal production systems.
In monogastric animals such as poultry and swine, a notable deficiency exists in the endogenous synthesis of enzymes necessary for the hydrolysis of non-starch polysaccharides (NSPs) like xylan (McLoughlin et al., 2017). This deficiency often manifests in poultry production as a decline in growth performance, attributed to increased digesta viscosity arising from the prevalence of NSPs in commonly utilized poultry feed ingredients. Without sufficient endogenous enzymes to degrade xylan, NSPs can increase digesta viscosity, encase essential nutrients, and create a barrier to their effective digestion. In response to this issue, monogastric animal producers have implemented exogenous enzymes such as xylanases into the feeds for swine and poultry to degrade xylan to short-chain sugars, thus reducing intestinal viscosity and improving the digestive utilization of nutrients (Sakata et al., 1995; Aragon et al., 2018)
Understanding Xylanase Enzymes
Xylanase enzymes belong to the class of carbohydrases that specifically target complex polysaccharides, such as xylan, a backbone nonstarch polysaccharide (NSP) prevalent in plant cell walls. These enzymes catalyze the hydrolysis of xylan into smaller, more digestible fragments, such as arabino–xylo-oligosaccharides (AXOs) and xylo-oligosaccharides (XOs), thereby facilitating the breakdown of dietary fiber in poultry diets.
Mechanism of action
It is generally agreed that the beneficial effects of feed xylanase are primarily due to the reduction in viscosity. Studies have shown that supplementing xylanases to animal feeds reduces digesta viscosity and releases encapsulated nutrients, thus improving the overall feed digestibility and nutrient availability (Matthiesen et al., 2021). The reduction in digesta viscosity by adding xylanase is achieved by the partial hydrolysis of NSPs in the upper digestive tract, leading to a decrease in digesta viscosity in the small intestine (Choct & Annison, 1992).
GH10 vs. GH11 Xylanases
Well-characterized xylanases are mostly grouped into glycoside hydrolase families 10 (GH10) and 11 (GH11) based on their structural characteristics (amino acid composition), mode of xylan degradation, the similarity of catalytic domains, substrate specificities, optimal conditions, thermostability, and practical applications.
Why are GH10 xylanases more efficient in animal production?
While both GH10 and GH11 xylanases act on the xylan main chain, these two enzyme types have different folds, substrate specificities, and mechanisms of action (Biely et al., 2016). The GH10 xylanases are more beneficial in animal feed production due to their efficient mechanism of action, broader substrate specificity, and better thermostability, as discussed below.
Generally, the GH10 xylanases exhibit broader substrate specificity and can hydrolyze various forms of xylan, including soluble and insoluble substrates. On the other hand, GH11 xylanases have a narrower substrate specificity and are primarily active on soluble xylan substrates. GH10 xylanases exhibit higher catalytic versatility and can catalyze the cleavage of the xylan backbone at the nonreducing side of substituted xylose residues, whereas GH11 enzymes require unsubstituted regions of the xylan backbone (Collins et al., 2005; Chakdar et al., 2016).
As a result, GH10 xylanases generally produce shorter xylo-oligosaccharides than members of the GH11 family (Collins et al., 2005). Moreover, as shown in Fig.1, the GH10 xylanase can rapidly and effectively break down xylan molecules.
Fig.1.: Activity of a bacterial GH10 xylanase against soluble and insoluble arabinoxylans
Higher thermostability
Enzymes are proteins, and the protein’s primary structure determines their thermostability. The enzyme protein tends to denature at higher than tolerable temperatures, rendering it inactive. An enzyme’s high-temperature tolerance ensures its efficacy throughout the pelleted feed manufacturing. This results in consistent enzyme activity in the finished feed, subsequent gut health, and predictable performance benefits.
Xylanases with higher thermostability are more suitable for applications requiring high-temperature processes. An intrinsically heat-stable bacterial xylanase maintains its activity even under high-temperature feed processing conditions, such as pelleting.
A study conducted at the University of Novi Sad, Serbia (Fig. 2), with three pelleting temperatures (85 °C, 90 °C, and 95 °C) and conditioning times of 4 and 6 mins, showed that Axxess XY, an intrinsically thermostable GH10 xylanase, demonstrated more than 85% recovery even at 4 to 6 mins conditioning time and 95 °C temperature.
Fig.2: Optimum recovery of Axxess XY at elevated conditioning time and temperatures
Maintaining consistently optimum enzyme activity is crucial for realizing the benefits of enzyme inclusion in feed under challenging feed processing conditions.
Conclusion
In conclusion, exogenous feed enzymes, including xylanase, have gained widespread recognition for their pivotal role in poultry nutrition. The increasing use of xylanase is attributed to its ability to effectively manage feed costs while incorporating high-fiber ingredients without compromising poultry performance. However, the efficacy of xylanase is based on several factors, including its mode of action, substrate specificity, catalytic efficacy, and thermostability. Selecting the appropriate xylanase enzyme tailored for specific needs is crucial to harnessing its full benefits.
A GH10 xylanase, such as Axxess XY, designed explicitly as a feed enzyme, offers distinct advantages in poultry production. Its efficient mechanism of action, broader substrate specificity, and superior thermostability make it a preferred choice for optimizing animal performance. Notably, Axxess XY exhibits exceptional activity against soluble and insoluble arabinoxylans, thereby enhancing nutrient utilization, promoting gut health, and ultimately elevating overall performance levels in poultry.
Incorporating specialized GH10 Xylanase enzymes like Axxess XY represents a strategic approach to unlocking the nutrients in feedstuffs, ensuring optimal performance, and maximizing profitability in the poultry business.
References
Aragon, Caio C., Ana I. Ruiz-Matute, Nieves Corzo, Rubens Monti, Jose M. Guisán, and Cesar Mateo. “Production of Xylo-Oligosaccharides (XOS) by Controlled Hydrolysis of Xylan Using Immobilized Xylanase from Aspergillus Niger with Improved Properties.” Integrative Food, Nutrition and Metabolism 5, no. 4 (2018). https://doi.org/10.15761/ifnm.1000225.
Bedford, Michael R., and Henry L. Classen. “Reduction of Intestinal Viscosity through Manipulation of Dietary Rye and Pentosanase Concentration Is Effected through Changes in the Carbohydrate Composition of the Intestinal Aqueous Phase and Results in Improved Growth Rate and Food Conversion Efficiency of Broiler Chicks.” The Journal of Nutrition 122, no. 3 (March 1992): 560–69. https://doi.org/10.1093/jn/122.3.560.
Biely, Peter, Suren Singh, and Vladimír Puchart. “Towards Enzymatic Breakdown of Complex Plant Xylan Structures: State of the Art.” Biotechnology Advances 34, no. 7 (November 2016): 1260–74. https://doi.org/10.1016/j.biotechadv.2016.09.001.
Choct, M., and G. Annison. “Anti‐nutritive Effect of Wheat Pentosans in Broiler Chickens: Roles of Viscosity and Gut Microflora.” British Poultry Science 33, no. 4 (September 1992): 821–34. https://doi.org/10.1080/00071669208417524.
Collins, Tony, Charles Gerday, and Georges Feller. “Xylanases, Xylanase Families and Extremophilic Xylanases.” FEMS Microbiology Reviews 29, no. 1 (January 2005): 3–23. https://doi.org/10.1016/j.femsre.2004.06.005.
Matthiesen, Connie F., Dan Pettersson, Adam Smith, Ninfa R. Pedersen, and Adam. C. Storm. “Exogenous Xylanase Improves Broiler Production Efficiency by Increasing Proximal Small Intestine Digestion of Crude Protein and Starch in Wheat-Based Diets of Various Viscosities.” Animal Feed Science and Technology 272 (February 2021): 114739. https://doi.org/10.1016/j.anifeedsci.2020.114739.
McLoughlin, Rebecca F, Bronwyn S Berthon, Megan E Jensen, Katherine J Baines, and Lisa G Wood. “Short-Chain Fatty Acids, Prebiotics, Synbiotics, and Systemic Inflammation: A Systematic Review and Meta-Analysis.” The American Journal of Clinical Nutrition 106, no. 3 (March 2017): 930–45. https://doi.org/10.3945/ajcn.117.156265.
Sakata, T., M. Adachi, M. Hashida, N. Sato, and T. Kojima. “Effect of N-Butyric Acid on Epithelial Cell Proliferation of Pig Colonic Mucosa in Short-Term Culture.” DTW – Deutsche Tierärztliche Wochenschau 102, no. 4 (1995): 163–64.
How xylanase can enhance swine productivity
By Dr. Ajay Awati, Director of Enzymes, EW Nutrition
The use of by-products and high-fiber ingredients in feed formulations has increased in swine operations. Driven by both economic and sustainability goals, this shift has emphasized the importance of understanding the role of dietary fibers and carbohydrases in swine nutrition and health (Petry & Patience, 2020). These feeds rich in fiber are generally considered to have low nutritional value due to the lower digestive energy or amino acid levels when compared to concentrated feeds with high starch or proteins (Woyengo et al., 2014).
Dietary fiber is vital in pig nutrition, necessitating a baseline inclusion to support regular digestive tract functions (Wenk, 2001). Incorporating fiber into the diets of monogastric animals raises concerns due to its correlation with reduced nutrient utilization and diminished net energy levels (Noblet; Le Goof, 2001). High-fiber diets can present challenges for inclusion in monogastric animals’ feeds, especially young animals, due to their bulky nature and restricted ability to ferment fiber, impacting nutrient uptake based on fiber type, the age of the pig, and diet composition (Bach Knudsen et al., 2012).
Moreover, the apparent ileal digestibility (AID) of nutrients is adversely affected by dietary fiber, attributed to the small intestine’s deficiency in endogenous enzymes necessary for breaking down these bonds (Bach Knudsen et al., 2012).
This article aims to demonstrate how enzymatic degradation of arabinoxylans, particularly through xylanase enzymes, can mitigate anti-nutritional effects and enhance the nutritional value of high-fiber swine diets, thereby improving animal health and performance.
Into the World of Arabinoxylans
In plants classified as monocotyledonous, such as cereals, the main non-starch polysaccharides (NSP’s) components of the cell wall are arabinoxylans, cellulose and β-glucan (Bach Knudsen, 1997). Arabinoxylans represent a complex group of dietary fibers with significant implications for swine nutrition and health. Their structural heterogeneity can influence physicochemical properties, biological activities, and affect pigs’ gut microbiota and immune system. Present in both soluble and insoluble forms, it consists of a backbone of xylose residues with arabinose side chains, playing a crucial role in the nutritional dynamics within swine diets (Mudgil & Barak, 2013).
The fermentability of corn-based dietary fiber is limited by its insoluble fraction and complex branched structure; impacting the digesta transit rate and reducing the digestibility of nutrients (Gutierrez et al., 2013). Supplementing exogenous carbohydrases offers a viable approach to enhance the utilization of fiber that is otherwise difficult to ferment, potentially amplifying its positive effects.
Xylanase’s Impact on Fiber and Gut Health
Non-digestible carbohydrates may be fermented by microbial populations along the gastrointestinal tract to synthesize short-chain fatty acids that may be absorbed and metabolized by the pig. Such indigestible carbohydrates consist of specific disaccharides, oligosaccharides, resistant starches, and non-starch polysaccharides. The presence and composition of these indigestible carbohydrates in pig diets vary based on the types of feed ingredients incorporated into their meals (Navarro et al., 2019). Xylanase works on the hydrolysis of the arabinoxylan fraction of NSPs. The NSPs present in the walls of plant cells encapsulate nutrients, making them unavailable for the action of the animal’s own digestive enzymes. Moreover, NSPs exhibit a high affinity for water within the gastrointestinal lumen, leading to elevated digesta viscosity. This increased viscosity reduces gastrointestinal motility, facilitating an environment conducive to the proliferation of pathogenic microflora (Choct, 1998). The advantageous outcomes of enzyme supplementation arise from the enzymatic disruption of intact cellular membranes, leading to the release of sequestered nutrients, or are a consequence of modifying the physicochemical properties of non-starch polysaccharides, due to changes in viscosity and water-holding capacity and/or changes in the composition and content of bacteria in the intestine (Bedford, M. R., & Classen, 1992).
Arabinoxylans in Cereal Grains and Their By-products
Factors such as genetics, climate, maturity stage, fertilizer use, and post-harvest storage time influence the proportion of total cell wall polysaccharides in cereal grains. These factors vary across production systems and countries, depending on the availability of feed resources (Paloheimo et al., 2010).
Cereal grains and their by-products, including wheat bran, corn distillers dried grains with solubles (DDGS), and rice husks, serve as significant sources of arabinoxylans. Their incorporation into swine diets is growing due to economic advantages.
The ethanol industry’s growth has increased the availability of distillery by-products. Brazil alone generates an estimated 366 million tons of DDGS annually (USDA, 2017). Among these by-products, distiller-dried grains are prevalent, especially in the U.S. pork industry as feed ingredients.
Corn, wheat, and barley, as staple ingredients in swine feed, exhibit significant variations in their NSP and arabinoxylans content. In grain form, corn contains 4.7% AX with a soluble component of 0.5%, while wheat has a higher arabinoxylans content at 7.3% with 1.8% being soluble. Barley stands out with the highest arabinoxylans content at 8.4%, of which 1.2% is soluble, reflecting its rich fiber composition. The processing into flour results in a reduction of arabinoxylans content across all three cereals, highlighting the impact of processing on dietary fiber availability (Knudsen, 2014).
Rice distillers’ by-product is recognized as a valuable protein source, boasting high crude protein levels. Nonetheless, its high fiber content can restrict usage (Huang et al., 2017). Wheat bran is particularly rich in arabinoxylans, enhancing its dietary fiber content. DDGS also contain significant amounts of both soluble and insoluble arabinoxylans, resulting from the corn kernel’s residual non-starch polysaccharides (Agyekum & Nyachoti, 2017).
It is essential to understand the specific levels of arabinoxylans in these components to create balanced diets that optimize nutritional benefits while minimizing potential anti-nutritional effects.
Addressing Arabinoxylan Degradation
Xylanases target specific substrates, necessitating the presence of arabinoxylans for their effective action. The complex structure of arabinoxylans makes them resistant to degradation by the swine’s endogenous enzymes, presenting a dual challenge: how to harness the beneficial effects of soluble arabinoxylans while mitigating the negative impacts of their insoluble counterparts.
These enzymes specifically cleave the 1,4-β-D-xylosidic bonds in arabinoxylans, randomly targeting xylose linkages within the xylan structure. Each enzyme type has a unique pattern of degradation (Collins et al., 2005) and GH 10 xylanases specialize in breaking down arabinoxylans with high arabinose substitution into smaller oligosaccharides. These oligosaccharides are valuable for fermentation, serving as energy sources or prebiotics.
Also, this group of enzymes action not only reduces gut viscosity but can lead to enhanced feed efficiency, growth performance, and overall health of swine by improving the digestibility of fibrous components in feed (Lærke et al., 2015). GH 10 xylanases often have optimal activity at pH levels found in the animal gut, and their thermal stability ensures they retain activity under feed processing temperatures. Lei et al. (2016) highlighted the efficacy of xylanase in improving nutrient digestibility and overall feed efficiency. By targeting the arabinoxylans present in swine diets, xylanase enzymes facilitate a more efficient conversion of feed into energy, contributing to improved growth rates and performance metrics.
As detailed by Tiwari, Singh, & Jha (2019), arabinoxylans undergo fermentation in the gut, leading to the production of short-chain fatty acids (SCFAs) that beneficially alter the gut microbial ecology. The application of GH 10 xylanases has been highlighted for its potential to significantly enhance the degradation of arabinoxylans, thereby improving the fermentation process and increasing the yield of SCFAs. This enzymatic breakdown facilitates more efficient nutrient absorption and overall better gastrointestinal health, directly influencing swine growth and performance positively.
A study reveals that xylanase supplementation significantly reduces mortality rates in pigs in a dose-dependent manner. With mortality rates dropping from 4.16% in the control group to as low as 2.25% with the highest xylanase dosage, the results highlight xylanase’s potential to improve gut health and increase survival rates. This suggests a promising approach for boosting pig well-being and reducing the reliance on enteric antibiotics, marking a significant stride in sustainable animal nutrition practices.(Zier-Rush et al., 2016).
The research conducted by Petry et al. (2020) demonstrated that xylanase increased the digestibility of non-starch polysaccharides, particularly arabinoxylan, in diets high in insoluble corn fiber. This improvement in nutrient absorption highlights xylanase’s role in optimizing the use of high-fiber ingredients in swine diets, thereby enhancing animal health and performance. Due to its cost-effectiveness and nutrient profile, xylanase supplementation enhances the nutritional value of DDG in swine diets.
The strategic implementation of xylanase in swine diets represents a promising approach to the challenges posed by high-fiber feed ingredients. By improving the digestibility of arabinoxylans and other complex carbohydrates, xylanase supplementation can mitigate the anti-nutritional effects of insoluble fibers, enhance feed efficiency, and support optimal growth and health outcomes in swine.
Enhancing Swine Productivity with Enzyme Solutions
With the growing incorporation of co-products in swine feed, there arises a crucial need to transform the high fiber content into a beneficial asset for the animals. The strategic incorporation of enzyme solutions, particularly xylanase enzymes, into swine feed formulations emerges as a scientifically supported approach to significantly enhance the digestibility of high-fiber diets. This method effectively addresses the nutritional intricacies posed by arabinoxylans, facilitating improved feed utilization. Moreover, the action of xylanase enzymes extends beyond enhancement of nutrient absorption; it plays a pivotal role in promoting the health and performance of swine. Such targeted nutritional strategies are vital in the context of swine production systems, highlighting the necessity of integrating these enzymatic solutions to achieve optimal animal health, growth, and productivity.
References:
Agyekum, K. A., & Nyachoti, C. M. (2017). Nutritional and metabolic consequences of feeding high-fiber diets to swine: A review. Engineering, 3(5), 716-725. https://doi.org/10.1016/J.ENG.2017.03.010
Bach Knudsen, K. E. (1997). Carbohydrate and lignin contents of plant materials used in animal feeding. Animal Feed Science and Technology, 67, 319-338.
Bach Knudsen, K. E., Hedemann, M. S., & Laerke, H. N. (2012). The role of carbohydrates in intestinal health of pigs. Animal Feed Science and Technology, 173, 41–53.
Bedford, M. R., & Classen, H. L. (1992). Reduction of intestinal viscosity through manipulation of dietary rye and pentosanase concentration is effected through changes in the carbohydrate composition of the intestinal aqueous phase and results in improved growth rate and food conversion efficiency of broiler chicks. The Journal of Nutrition, 122, 560-569.
Choct, M. (1998). The effect of different xylanases on carbohydrate digestion and viscosity along the intestinal tract in broilers. Australian Poultry Science Symposium, 10.
Collins, T., Gerday, C., & Feller, G. (2005). Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiology Reviews, 29(1), 3–23. https://doi.org/10.1016/j.femsre.2004.06.005
Gutierrez, N. A., Kerr, B. J., & Patience, J. F. (2013). Effect of insoluble-low fermentable fiber from corn-ethanol distillation origin on energy, fiber, and amino acid digestibility, hindgut degradability of fiber, and growth performance of pigs. Journal of Animal Science, 91, 5314–5325. https://doi.org/10.2527/jas.2013-6328
Huang, Y. F., Gao, X. L., Nan, Z. B., & Zhang, Z. X. (2017). Potential value of the common vetch (Vicia sativa L.) as an animal feedstuff: A review. Journal of Animal Physiology and Animal Nutrition, 101, 807-823. https://doi.org/10.1111/jpn.12617
Lærke, H. N., Arent, S., Dalsgaard, S., & Bach Knudsen, K. E. (2015). Effect of xylanases on ileal viscosity, intestinal fiber modification, and apparent ileal fiber and nutrient digestibility of rye and wheat in growing pigs. Journal of Animal Science, 93(9), 4323-4335.
Lei, Z., Shao, Y., Yin, X., Yin, D., Guo, Y., & Yuan, J. (2016). Combination of xylanase and debranching enzymes specific to wheat arabinoxylan improve the growth performance and gut health of broilers. Journal of Agricultural and Food Chemistry, 64(24), 4932-4942.
Mudgil, D., & Barak, S. (2013). Composition, properties and health benefits of indigestible carbohydrate polymers as dietary fiber: a review. International Journal of Biological Macromolecules, 61, 1-6. https://doi.org/10.1016/j.ijbiomac.2013.06.044
Navarro, D. M. D. L., Abelilla, J. J., & Stein, H. H. (2019). Structures and characteristics of carbohydrates in diets fed to pigs: a review. Journal of Animal Science and Biotechnology, 10, 39. https://doi.org/10.1186/s40104-019-0345-6
Noblet, J., & Le Goff, G. I. (2001). Effect of dietary fibre on the energy value of feeds for pigs. Animal Feed Science and Technology, 90, 35-52.
Paloheimo, M., Piironen, J., & Vehmaanperä, J. (2010). Xylanases and cellulases as feed additives. In M. Bedford & G. Partridge (Eds.), Enzymes in farm animal nutrition (2nd ed., pp. 12-53). CABI Publishing. https://doi.org/10.1079/9781845936747.0012
Petry, A., Huntley, N., Bedford, M., Zijlstra, R. T., & Patience, J. (2020). Supplementing xylanase increased the digestibility of non-starch polysaccharides, particularly arabinoxylan, in diets high in insoluble corn fiber fed to swine with a 36-d dietary adaptation period. Journal of Animal Science, 98(52-52).
Petry, A. L., & Patience, J. F. (2020). Xylanase supplementation in corn-based swine diets: a review with emphasis on potential mechanisms of action. Journal of Animal Science, 98(11), skaa318. https://doi.org/10.1093/jas/skaa318
Tiwari, U., Singh, A., & Jha, R. (2019). Fermentation characteristics of resistant starch, arabinoxylan, and β-glucan and their effects on the gut microbial ecology of pigs: A review. Animal Nutrition, 5, 217-226.
United States Department of Agriculture (USDA). (2017). Biofuel Annual. Economic Research Service. https://www.fas.usda.gov/commodities/biofuels
Wenk, C. (2001). The role of dietary fibre in the digestive physiology of the pig. Animal Feed Science and Technology, 90, 21-33.
Woyengo, T. A., Beltranena, E., & Zijlstra, R. T. (2014). Nonruminant nutrition symposium: controlling feed cost by including alternative ingredients into pig diets: a review. Journal of Animal Science, 92(4), 1293-1305.
Zier-Rush, C. E., Groom, C., Tillman, M., Remus, J., & Boyd, R. D. (2016). The feed enzyme xylanase improves finish pig viability and carcass feed efficiency. Journal of Animal Science, 94(suppl_2), 115. https://doi.org/10.2527/msasas2016-244
Overcoming Challenges of Xylanase Inhibitors in Animal Feeds
By Dr. Ajay Awati, Global Director Enzymes, EW Nutrition
In recent years, the scientific understanding of xylanase inhibitors (XIs) and their impact on animal nutrition has grown significantly. Xylanase, a crucial enzyme used to enhance nutrient availability in feed, can face challenges from XIs present in cereal grains. This article explores the evolution of plant protection mechanisms, the economic impact of XIs, and the development of a novel xylanase, Axxess XY, resistant to these inhibitors.
Xylanase inhibitors – an evolutionary protection mechanism of plants
Xylanase inhibitors (XI) are a classic example of the evolutionary development of protection mechanisms by cereal plants against pathogens. Microorganisms, such as fungal pathogens, involve the degradation of xylan as one of the mechanisms in pathogenesis (Choquer et al., 2007). There are also other mechanisms by which microorganism-produced xylanases affect plants.
To protect themselves, plants evolved xylanase inhibitors to prevent the activities of xylanases. XIs are plant cell wall proteins broadly distributed in monocots. There are three classes of XIs with different structures and inhibition specificities (Tundo et al., 2022):
1. Triticum aestivum xylanase inhibitors (TAXI)
2. Xylanase inhibitor proteins (XIP), and
3. Thaumatin-like xylanase inhibitors (TLXI).
Xylanase inhibitors have an economic impact
In animal nutrition, xylanases are widely used in diets containing cereal grains and other plant materials to achieve a higher availability of nutrients. The inhibitory activity of XIs prevents this positive effect of the enzymes and, therefore, makes them economically relevant. Studies have reported that higher levels of XIs negatively impact broiler performance. For example, in one of the studies, broilers fed with grains of a cultivar with high inhibitory activity showed a 7% lower weight on day 14 than broilers fed with grains of a cultivar with less inhibitory activity (Madesen et al., 2018). Another study by Ponte et al. (2004) also concluded that durum wheat xylanase inhibitors reduced the activity of exogenous xylanase added to the broiler diets.
Xylanase inhibitors can withstand high temperatures
Even though XIs can impact the performance of exogenous xylanase in different ways, only minor attention was paid to the reduction of xylanase’s susceptibility to xylanase inhibitors during the xylanase development in the last decades. Firstly, the issue was ignored mainly through the assumption that XIs are denatured or destroyed during pelleting processes. However, Smeets et al. (2014) showed that XIs could sustain significant temperature challenges. They demonstrated that after exposing wheat to pelleting temperatures of 80°C, 85°C, 92°C, and 95°C, the recovery of inhibitory activity was still 99%, 100%, 75%, and 54%, respectively. Furthermore, other studies also confirmed that conditioning feed at 70-90°C for 30 sec followed by pelleting had little effect on the XI activity in the tested feed, showing that xylanase inhibitors are very likely present in most xylanase-supplemented feeds fed to animals.
Do we only have the problem of xylanase inhibitors in wheat?
No. After first reports of the presence of xylanase inhibitors in wheat by Debyser et al. (1997, 1999), XIs were also found in other cereal grains (corn, rice, and sorghum, etc.), and their involvement in xylanase inhibition and plant defense has been established by several reports (Tundo et al., 2022).
In most of the countries outside Europe, exogenous xylanase is used not only in wheat but also in corn-based diets. Besides broiler feeds, also other animal feeds, such as layer or swine feed being part of more mixed-grain diets, are susceptible to the inhibitory activity of XIs. Nowadays, the situation is getting worse with all the raw material prices increasing and nutritionists tending to use other feed ingredients and locally produced cereals. They need a xylanase which is resistant to xylanase inhibitors.
Xylanases’ resistance to XIs is crucial – Axxess XY shows it
To prevent xylanases from losing their effect due to the presence of xylanase inhibitors, the resistance of new-generation xylanases to these substances is paramount in the development process, including enzyme discovery and engineering.
In the past 25 years, scientists have learned much about XI-encoding genes and discovered how xylanase inhibitors can block microbial xylanases. Additionally, there has been a significant increase in understanding the structural aspects of the interaction between xylanases and XIs, mainly how xylanase inhibitors interact with specific xylanases from fungi or bacteria and those in the GH10 or GH11 family. With such understanding, a new generation xylanase, Axxess XY, was developed. Besides showing the essential characteristics of intrinsic thermostability and versatile activity on both soluble and insoluble arabinoxylan, it is resistant to xylanase inhibitors.
Axxess XY takes xylanase application in animal feeds to the next level.
Axxess XY outperforms other xylanases on the market
Recent scientific developments (Fierens, 2007; Flatman et al., 2002; Debyser, 1999; Tundo et al., 2022; Chmelova, 2019) and internal research can be summarized as follows:
Figure 1: Schematic summary of the susceptibility of different xylanase to xylanase inhibitors from three main groups.
The high resistance to xylanase inhibitors is one of the reasons that a novel xylanase with bacterial origin and from the GH-10 family was chosen to be Axxess XY. EWN innovation, together with research partners, made an interesting benchmark comparison between xylanases that are commercially sold by different global suppliers and Axxess XY. For these trials, all xylanase inhibitors from wheat were extracted. The inhibitors, together with the respective xylanase, were incubated at 400C (to mimic birds’ body temperature) for 30 mins. Then, the loss of xylanase activity was calculated by analyzing remaining activity after incubation. Results are shown below in Figure 2. There were varying levels of activity loss observed in the different commercially sold xylanases. In some xylanases, the losses were alarmingly high. However, Axxess XY was not inhibited at all.
Fig. 2: Extracted total xylanase inhibitors from wheat incubated with the respective xylanase at 40°C for 30 mins. – Loss of activity after incubation with xylanase inhibitors
Conclusion:
Xylanase inhibitors are present in all cereal grains and, unfortunately, heat tolerant (up to 900C, still 75% of inhibition activity was retained). Regardless of the diets used, there is a possibility that the xylanase used may come across xylanase inhibitors, resulting in a loss of activity. More importantly, this can lead to inconsistent performance.
For effective, consistent, and higher performance of NSP enzyme application, it is a must to use xylanase that is resistant to xylanase inhibitors.
Literature:
Chmelová, Daniela, Dominika Škulcová, and Miroslav Ondrejovic. “Microbial Xylanases and Their Inhibition by Specific Proteins in Cereals.” KVASNY PRUMYSL 65, no. 4 (2019). https://doi.org/10.18832/kp2019.65.127. LINK
Choquer, Mathias, Elisabeth Fournier, Caroline Kunz, Caroline Levis, Jean-Marc Pradier, Adeline Simon, and Muriel Viaud. “Botrytis CinereaVirulence Factors: New Insights into a Necrotrophic and Polyphageous Pathogen.” FEMS Microbiology Letters 277, no. 1 (2007): 1–10. https://doi.org/10.1111/j.1574-6968.2007.00930.x. LINK
Debyser, W, WJ Peumans, EJM Van Damme, and JA Delcour. “Triticum Aestivum Xylanase Inhibitor (Taxi), a New Class of Enzyme Inhibitor Affecting Breadmaking Performance.” Journal of Cereal Science 30, no. 1 (1999): 39–43. https://doi.org/10.1006/jcrs.1999.0272. LINK
Organic acids can play a crucial role in zinc oxide replacement
Dr. Inge Heinzl, Editor EW Nutrition & Juan Antonio Mesonero Escuredo, GTM Swine/GPM Organic Acids EW Nutrition
The use of high levels of Zinc Oxide (ZnO) in the EU before 2022 was one of the most common methods to prevent postweaning diarrhea (PWD) in pig production. Pharmacologically high levels of ZnO (2000-3000 ppm) increase growth and reduce the incidence of enteric bacterial diseases such as post-weaning diarrhea (PWD)( Carlson et al., 1999; Hill et al., 2000; Hill et al., 2001; Poulsen & Larsen, 1995; De Mille et al., 2019).
However, ZnO showed adverse effects, such as the accumulation of heavy metal in the environment, the risk for antimicrobial resistance (AMR), and problems of mineral toxicity and adverse growth effects when feeding it longer than 28 days (Jensen et al., 2018; Cavaco et al., 2011; Vahjen, 2015; Romeo et al., 2014; Burrough et al., 2019). To replace ZnO in pig production, let us first look at its positive effects to know what we must compensate for.
ZnO has a multifactorial mode of action
ZnO shows several beneficial characteristics that positively influence gut health, the immune system, digestion, and, therefore, also overall health and growth performance.
Figure 1. Beneficial effects and ZnO mode of action in postweaning piglets
1. ZnO acts as an antimicrobial
Concerning the antimicrobial effects of ZnO, different possible modes of action are discussed:
ZnO in high dosages generates reactive oxygen species (ROS) that can damage the bacterial cell walls (Pasquet et al., 2014)
The death of the bacterial cell due to direct contact of the metallic Zn to the cell (Shearier et al., 2016)
Intrinsic antimicrobial properties of the ZnO2+ ions after dissociation. The uptake of zinc into cells is regulated by homeostasis. A concentration of the ZnO2+ ions higher than the optimal level of 10-7 to 10-5 M (depending on the microbial strain) allows the invasion of Zn2+ ions into the cell, and the zinc starts to be cytotoxic (Sugarman, 1983; Borovanský et al., 1989).
ZnO shows activity against, e.g., Staphylococcus aureus, Pseudomonas aeruginosa, E. coli,Streptococcus pyogenes, and other enterobacteria (Ann et al., 2014; Vahjen et al., 2016). However, Roselli et al. (2003) did not see a viability-decreasing effect of ZnO on ETEC.
2. ZnO modulates the immune system
Besides fighting pathogenic organisms as described in the previous chapter and supporting the immune system, ZnO is an essential trace element and has a vital role in the immune system. ZnO improves the innate immune response, increasing phagocytosis and oxidative bursts from macrophages and neutrophils. It also ameliorates the adaptative immune response by increasing the number of T lymphocytes (T cells) in general and regulatory T lymphocytes (T-regs) in particular. These cells control the immune response and inflammation (Kloubert et al., 2018). Macrophage capacity for phagocytosis (Ercan and Bor, 1991) and to kill parasites (Wirth et al., 1989), and also the killing activity of natural killer cells depends on Zn (Rolles et al., 2018). By reducing bacterial adhesion and blocking bacterial invasion, ZnO disburdens the immune system (Roselli et al., 2003).
ZnO reduces the expression of several proinflammatory cytokines induced by ETEC (Roselli et al., 2003). Several studies have also shown a modulation effect on intestinal inflammation, decreasing levels of IFN-γ, TNF-α, IL-1ß and IL-6, all pro-inflammatory, in piglets supplemented with ZnO (Zhu et al., 2017; Grilli et al., 2015).
3. ZnO improves digestion and promotes growth
Besides protecting young piglets against diarrhea, the goal is to make them grow optimally. For this target, an efficient digestion and a high absorption of nutrients is essential. Stimulating diverse pancreatic enzymes such as amylase, carboxypeptidase A, trypsin, chymotrypsin, and lipase increases digestibility (Hedemann et al., 2006; Pieper et al., 2015). However, Pieper et al. (2015) also showed that a long-term supply of very high dietary zinc triggers oxidative stress in the pancreas of piglets.
By stimulating the secretion of ghrelin at the stomach level and thereby promoting the release of insulin-like growth factor (IGF-1) and cholecystokinin (CCK), ZnO enhances muscle protein synthesis, cell proliferation, and feed intake (Yin et al., 2009; MacDonald et al., 2000)).
The result of improved digestion is increased body weight and average daily gain, which can be seen, e.g., in a study by Zhu et al. (2017).
4. ZnO protects the intestinal morphology
ZnO prevents the decrease of the trans-endothelial electrical resistance (TEER), usually occurring in the case of inflammation, by downregulating TNF-α and IFN-γ. TNF-α, as well as IFN-γ, increase the permeability of the epithelial tight junctions and, therefore, the intestinal barrier (Al-Sadi et al., 2009).
The enterotrophic and anti-apoptotic effect of ZnO is reflected by a higher number of proliferating and PCNA-positive cells and an increased mucosa surface in the ileum (higher villi, higher villi/crypt ratio)(Grilli et al., 2015). Zhu et al. (2017) also saw an increase in villus height in the duodenum and ileum and a decrease in crypt depth in the duodenum due to the application of 3000 mg of ZnO/kg. Additionally, they could notice a significant (P<0.05) upregulation of the mRNA expression of the zonula occludens-1 and occluding in the mucosa of the jejunum of weaned piglets.
In a trial conducted by Roselli et al. (2003), the supplementation of 0.2 mmol/L ZnO prevented the disruption of the membrane integrity when human Caco-2 enterocytes were challenged with ETEC.
5. ZnO acts antioxidant
The antioxidant effect of ZnO was shown in a study conducted by Zhu et al., 2017. They could demonstrate that the concentration of malondialdehyde (MDA), a marker for lipid peroxidation, decreased on day 14 or 28, and the total concentration of superoxide dismutase (SOD), comprising enzymes that transform harmful superoxide anions into hydrogen peroxide, increased on day 14 (P<0.05). Additionally, Zn is an essential ion for the catalytic action of these enzymes.
Which positive effects of ZnO can be covered by organic acids (OAs)?
1. OAs act antimicrobial
OAs, on the one hand, lower the pH in the gastrointestinal tract. Some pathogenic bacteria are susceptible to low pH. At a pH<5, the proliferation of, e.g., Salmonella, E. coli, and Clostridium is minimized. The good thing is that some beneficial bacteria, such as lactobacilli or bifidobacteria, survive as they are acid-tolerant. The lactobacilli, on their side, can produce hydrogen peroxide, which inhibits, e.g., Staphylococcus aureus or Pseudomonas spp. (Juven and Pierson, 1996).
Besides this more indirect mode of action, a more direct one is also possible: Owing to their lipophilic character, the undissociated form of OAs can pass the bacterial membrane (Partanen and Mroz, 1999). The lower the external pH, the more undissociated acid is available for invading the microbial cells. Inside the cell, the pH is higher than outside, and the OA dissociates. The release of hydrogen ions leads to a decrease in the internal pH of the cell and to a depressed cell metabolism. To get back to “normal conditions”, the cell expels protons. However, this is an energy-consuming process; longer exposure to OAs leads to cell death. The anion remaining in the cell, when removing the protons, disturbs the cell’s metabolic processes and participates in killing the bacterium.
These theoretical effects could be shown in a practical trial by Ahmed et al. (2014). He fed citric acid (0.5 %) and a blend of acidifiers composed of formic, propionic, lactic, and phosphoric acid + SiO2 (0.4 %) and saw a reduction in fecal counts of Salmonella and E. coli for both groups.
2. OAs modulate the immune system
The immune system is essential in the pig’s life, especially around weaning. Organic acids have been shown to support or stimulate the immune system. Citric acid (0.5%), as well as the blend of acidifiers mentioned before (Ahmed et al., 2014), significantly increased the level of serum IgG. IgG is part of the humoral immune system. They mark foreign substances to be eliminated by other defense systems.
Ren et al. (2019) could demonstrate a decrease in plasma tumor necrosis factor-α that regulates the activity of diverse immune cells. He also found lower interferon-γ and interleukin (Il)-1ß values in the OA group than in the control group after the challenge with ETEC. This trial shows that inflammatory response can be mitigated through the addition of organic acids.
3. OAs improve digestion and promote growth
In piglets, the acidity in the stomach is responsible for the activation and stimulation of certain enzymes. Additionally, it keeps the feed in the stomach for a longer time. Both effects lead to better digestion of the feed.
In the stomach, the conversion of pepsinogen to pepsin, which is responsible for protein digestion, is catalyzed under acid conditions (Sanny et al., 1975)group. Pepsin works optimally at two pH levels: pH 2 and pH 3.5 (Taylor, 1959). With increasing pH, the activity decreases; at pH 6, it stops. Therefore, a high pH can lead to poor digestion and undigested protein arriving in the intestine.
These final products of pepsin protein digestion are needed in the lower parts of the GIT to stimulate the secretion of pancreatic proteolytic enzymes. If they do not arrive, the enzymes are not activated, and the inadequate protein digestion continues. Additionally, gastric acid is the primary stimulant for bicarbonate secretion in the pancreas, neutralizing gastric acid and providing an optimal pH environment for the digestive enzymes working in the duodenum.
As already mentioned, the pH in the stomach influences the transport of digesta. The amount of digesta being transferred from the stomach to the small intestine is related to the acidity of the chyme leaving the stomach and arriving in the small intestine. Emptying of the stomach can only take place when the duodenal chyme can be neutralized by pancreatic or other secretions (Pohl et al., 2008); so, acid-sensitive receptors provide feedback regulation and a higher pH in the stomach leads to a faster transport of the digesta and a worse feed digestion.
4. OAs protect the intestinal morphology
Maintaining an intact gut mucosa with a high surface area is crucial for optimal nutrient absorption. Research suggests organic acids play a significant role in improving mucosal health:
Butyric acid promotes epithelial cell proliferation, as demonstrated in an in vitro pig hindgut mucosa study (Sakata et al., 1995). Fumaric acid, serving as an energy source, may locally enhance small intestinal mucosal growth, aiding in post-weaning epithelial cells’ recovery and increasing absorptive surface and digestive capacity (Blank et al., 1999). Sodium butyrate supplementation at low doses influences gastric morphology and function, thickening the stomach mucosa and enhancing mucosal maturation and differentiation (Mazzoni et al., 2008).
Studies show that organic acids affect gut morphology, with a mixture of short-chain and mid-chain fatty acids leading to longer villi (Ferrara et al., 2016) and Na-butyrate supplementation increasing crypt depth and villi length in the distal jejunum and ileum (Kotunia et al., 2004). However, the villi length and mucosa thickness in the duodenum were reduced. Dietary sodium butyrate has been linked to increased microvilli length and cecal crypt depth in pigs (Gálfi and Bokori, 1990).
5. OAs show antioxidant activity
The last characteristic, the antioxidant effect, cannot be provided at the same level as with ZnO; however, Zhang et al. (2019) attest to OAs a certain antioxidant activity. Oxalic, citric, acetic, malic, and succinic acids, which were extracted from Camellia oleifera, also showed good antioxidant activity in a trial conducted by Zhang et al. (2020).
Organic acids are an excellent tool to compensate for the ban on ZnO
The article shows that organic acids have similar positive effects as zinc oxide. They act antimicrobial, modulate the immune system, maintain the gut morphology, fight pathogenic microbes, and also act – slightly – antioxidant. Additionally, they have a significant advantage: they are not harmful to the environment. Organic acids used in the proper pH range and combination are good tools for replacing zinc oxide.